
Patterns of Endemism in Lichens: Another Paradigm-Shifting Example in the Lichen Genus Xanthoparmelia from Macaronesia
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Morphology and Anatomy
2.2. Molecular Analysis
2.2.1. DNA Isolation, Amplification, and Sequencing
2.2.2. Sequence Alignment and Phylogenetic Analysis
2.3. Dating Analysis
2.4. Macroclimatic Niche Characterization
3. Results
3.1. Sequence Alignment and Phylogenetic Analysis
3.2. Dating Analysis
3.3. Macroclimatic Niche Characterization
3.4. Taxonomy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nash, T.H., III. Nitrogen, its metabolism and potential contribution to ecosystems. In Lichen Biology; Cambridge University Press: Cambridge, UK, 2008; pp. 216–233. [Google Scholar]
- Honegger, R. Lichen-forming fungi and their photobionts. In Plant Relationships; Springer: Berlin/Heidelberg, Germany, 2009; pp. 307–333. [Google Scholar]
- Sanders, W.B. The disadvantages of current proposals to redefine lichens. New Phytol. 2023, 241, 969–971. [Google Scholar] [CrossRef]
- Galloway, D.J. Lichen biogeography. In Lichen Biology, 2nd ed.; Nash, T.H., Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 315–335. ISBN 978-0-521-69216-8. [Google Scholar]
- Fontaneto, D. Biogeography of Microscopic Organisms: Is Everything Small Everywhere? Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Muñoz, J.; Felicísimo, Á.M.; Cabezas, F.; Burgaz, A.R.; Martínez, I. Wind as a long-distance dispersal vehicle in the Southern Hemisphere. Science 2004, 304, 1144–1147. [Google Scholar] [CrossRef]
- Werth, S.; Wagner, H.H.; Gugerli, F.; Holderegger, R.; Csencsics, D.; Kalwij, J.M.; Scheidegger, C. Quantifying dispersal and establishment limitation in a population of an epiphytic lichen. Ecology 2006, 87, 2037–2046. [Google Scholar] [CrossRef]
- Damialis, A.; Kaimakamis, E.; Konoglou, M.; Akritidis, I.; Traidl-Hoffmann, C.; Gioulekas, D. Estimating the abundance of airborne pollen and fungal spores at variable elevations using an aircraft: How high can they fly? Sci. Rep. 2017, 7, 44535. [Google Scholar] [CrossRef] [PubMed]
- Scheidegger, C.; Werth, S. Conservation strategies for lichens: Insights from population biology. Fungal Biol. Rev. 2009, 23, 55–66. [Google Scholar] [CrossRef]
- Golan, J.J.; Pringle, A. Long-distance dispersal of fungi. Microbiol. Spectr. 2017, 5, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Werth, S. Biogeography and phylogeography of lichen fungi and their photobionts. In Biogeography of Microscopic Organisms. Is Everything Small Everywhere? Fontaneto, D., Ed.; Cambridge University Press: Cambridge, UK, 2011; pp. 191–208. ISBN 978-0-521-76670-8. [Google Scholar]
- Serusiaux, E.; Villarreal, A.J.C.; Wheeler, T.; Goffinet, B. Recent origin, active speciation and dispersal for the lichen genus Nephroma (Peltigerales) in Macaronesia. J. Biogeogr. 2011, 38, 1138–1151. [Google Scholar] [CrossRef]
- Moncada, B.; Reidy, B.; Lücking, R. A phylogenetic revision of Hawaiian Pseudocyphellaria sensu lato (lichenized Ascomycota: Lobariaceae) reveals eight new species and a high degree of inferred endemism. Bryol. 2014, 117, 119–160. [Google Scholar] [CrossRef]
- Lücking, R.; Dal-Forno, M.; Sikaroodi, M.; Gillevet, P.M.; Bungartz, F.; Moncada, B.; Yánez-Ayabaca, A.; Chaves, J.L.; Coca, L.F.; Lawrey, J.D. A single macrolichen constitutes hundreds of unrecognized species. Proc. Natl. Acad. Sci. USA 2014, 111, 11091–11096. [Google Scholar] [CrossRef] [PubMed]
- Perez-Vargas, I.; Perez-Ortega, S. A new endemic Ramalina species from the Canary Islands (Ascomycota, Lecanorales). Phytotaxa 2014, 159, 269–278. [Google Scholar] [CrossRef]
- Simon, A.; Goffinet, B.; Magain, N.; Sérusiaux, E. High diversity, high insular endemism and recent origin in the lichen genus Sticta (lichenized Ascomycota, Peltigerales) in Madagascar and the Mascarenes. Mol. Phylogenet. Evol. 2018, 122, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Benavent, I.; de Los Ríos, A.; Núñez-Zapata, J.; Ortiz-Álvarez, R.; Schultz, M.; Pérez-Ortega, S. Ocean crossers: A tale of disjunctions and speciation in the dwarf-fruticose Lichina (lichenized Ascomycota). Mol. Phylogenet. Evol. 2023, 185, 107829. [Google Scholar] [CrossRef] [PubMed]
- Ertz, D.; Tehler, A. New species of Arthoniales from Cape Verde with an enlarged concept of the genus Ingaderia. Lichenologist 2023, 55, 1–15. [Google Scholar] [CrossRef]
- Jorna, J.; Linde, J.B.; Searle, P.C.; Jackson, A.C.; Nielsen, M.E.; Nate, M.S.; Saxton, N.A.; Grewe, F.; de los Angeles Herrera-Campos, M.; Spjut, R.W.; et al. Species boundaries in the messy middle—A genome-scale validation of species delimitation in a recently diverged lineage of coastal fog desert lichen fungi. Ecol. Evol. 2021, 11, 18615–18632. [Google Scholar] [CrossRef]
- Ortiz-Álvarez, R.; de Los Ríos, A.; Fernández-Mendoza, F.; Torralba-Burrial, A.; Pérez-Ortega, S. Ecological specialization of two photobiont-specific maritime cyanolichen species of the genus Lichina. PLoS ONE 2015, 10, e0132718. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Palacios, J.M.; de Nascimento, L.; Otto, R.; Delgado, J.D.; García-del-Rey, E.; Arévalo, J.R.; Whittaker, R.J. A reconstruction of Palaeo-Macaronesia, with particular reference to the long-term biogeography of the Atlantic island laurel forests. J. Biogeogr. 2011, 38, 226–246. [Google Scholar] [CrossRef]
- Juan, C.; Emerson, B.C.; Oromí, P.; Hewitt, G.M. Colonization and diversification: Towards a phylogeographic synthesis for the Canary Islands. Trends Ecol. Evol. 2000, 15, 104–109. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Conservation International 2023. Available online: https://www.cepf.net/node/1996 (accessed on 1 December 2023).
- Medail, F.; Quezel, P. Hot-spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Medail, F.; Quezel, P. Biodiversity hotspots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Pérez-Vargas, I. A revised checklist of the lichen and lichenicolous fungi from the Canary Islands. In prep.
- García-Romero, L.; Carreira-Galbán, T.; Rodríguez-Báez, J.A.; Máyer-Suárez, P.; Hernández-Calvento, L.; Yánes-Luque, A. Mapping environmental impacts on coastal tourist áreas of oceanic islands (Gran Canaria, Canary Islands): A current and future scenarios assessment. Remote Sens. 2023, 15, 1586. [Google Scholar] [CrossRef]
- Hansen Machín, A.R.; Moreno Medina, C.J.; Naranjo Cigala, A.; Pérez Torrado, F.J.; Ramos, A.; Salas Pascual, M. Geografía de un islote: La Isleta de Gran Canaria. In Canarias. Los Valores Naturales de las Propiedades del Minsiterio de Defensa; Anadón Fernandez, I., Coord., Eds.; Ministerio de Defensa, Secretaría General Técnica: Madrid, Spain, 2021; pp. 106–149. [Google Scholar]
- Pérez-Vargas, I. (Department of Botany, Ecology and Plant Physiology, Faculty of Pharmacy, University of La Laguna, Apdo Postal 456, 38200 La Laguna, Canary Islands, Spain). Servicio de Evaluación y Caracterización de la Biota Liquénica del Paisaje Protegido de La Isleta. Gran Canaria. Informe Técnico, Tenerife. 2020; Unpublished work. [Google Scholar]
- Blanco, O.; Crespo, A.; Elix, J.A.; Hawksworth, D.L.; Lumbsch, H.T. A molecular phylogeny and a new classification of parmelioid lichens containing Xanthoparmelia-type lichenan (Ascomycota: Lecanorales). Taxon 2004, 53, 959–975. [Google Scholar] [CrossRef]
- Jaklitsch, W.M.; Baral, H.O.; Lücking, R.; Lumbsch, H.T. Syllabus of Plant Families—Adolf Engler’s Syllabus Der Pflanzenfamilien; Borntraeger Verlagsbuchhandlung: Stuttgart, Germany, 2016. [Google Scholar]
- Lücking, R.; Hodkinson, B.P.; Leavitt, S.D. The 2016 classification of lichenized fungi in the Ascomycota and Basidiomycota–Approaching one thousand genera. Bryologist 2017, 119, 361–416. [Google Scholar] [CrossRef]
- Leavitt, S.D.; Kirika, P.M.; Paz, G.A.D.; Huang, J.-P.; Hur, J.-S.; Elix, J.A.; Grewe, F.; Divakar, P.K.; Lumbsch, H.T. Assessing phylogeny and historical biogeography of the largest genus of lichen-forming fungi, Xanthoparmelia (Parmeliaceae, Ascomycota). Lichenol. 2018, 50, 299–312. [Google Scholar] [CrossRef]
- Hale, M.E., Jr. A synopsis of the lichen genus Xanthoparmelia (Vainio) Hale (Ascomycotina, Parmeliaceae). Smithson. Contrib. Bot. 1990, 74, 1–250. [Google Scholar] [CrossRef]
- Blanco, O.; Crespo, A.; Ree, R.H.; Lumbsch, H.T. Major clades of parmelioid lichens (Parmeliaceae, Ascomycota) and the evolution of their morphological and chemical diversity. Mol. Phylogenet. Evol. 2006, 39, 52–69. [Google Scholar] [CrossRef] [PubMed]
- Rizzi, G.; Giordani, P. The ecology of the lichen genus Xanthoparmelia in Italy: An investigation throughout spatial scales. Plant Biosyst. 2013, 147, 33–39. [Google Scholar] [CrossRef]
- Blanco, O.; Crespo, A.; Elix, J.A. Two new species of Xanthoparmelia (Ascomycota: Parmeliaceae) from Spain. Lichenologist 2005, 37, 97–100. [Google Scholar] [CrossRef]
- Crespo, A.; Kauff, F.; Divakar, P.K.; del Prado, R.; Pérez-Ortega, S.; Amo de Paz, G.; Ferencova, Z.; Blanco, O.; Roca-Valiente, B.; Núñez-Zapata, J.; et al. Phylogenetic generic classification of parmelioid lichens (Parmeliaceae, Ascomycota) based on molecular, morphological and chemical evidence. Taxon 2010, 59, 1735–1753. [Google Scholar] [CrossRef]
- Matteucci, E.; Occhipinti, A.; Piervittori, R.; Maffei, M.; Favero-Longo, S.E. Morpho-Chemical Variability of Xanthoparmelia (Vain.) Hale at the Local Scale: Unexpected Patterns and Edaphic Influence; IAL 8, Book of Abstracts; International Association for Lichenology: Helsinki, Finland, 2016; p. 117. [Google Scholar]
- Orange, A.; James, P.W.; White, F.J. Microchemical Methods for the Identification of Lichens; British Lichen Society: London, UK, 2001. [Google Scholar]
- Elix, J.A.; Ernst-Russell, K.D. A Catalogue of Standardized Thin Layer Chromatographic Data and Biosynthetic Relationships for Lichen Substances; Australian National University, Department of Chemistry: Canberra, Australia, 1993. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes—Application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.D.; Lee, S.; Taylor, J. Application and direct sequencing of fungal ribosomal DNA for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Mangold, A.; Martín, M.P.; Kalb, K.; Lücking, R.; Lumbsch, H.T. Molecular data show that Topeliopsis (Ascomycota, Thelotremataceae) is polyphyletic. Lichenologist 2008, 40, 39–46. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified risbosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4239–4246. [Google Scholar] [CrossRef] [PubMed]
- Zoller, S.; Scheidegger, C.; Sperisen, C. PCR primers for the amplification of mitochondrial small subunit ribosomal DNA of lichen-forming ascomycetes. Lichenologist 1999, 31, 511–516. [Google Scholar] [CrossRef]
- Zhou, S.; Stanosz, G.R. Primers for amplification of mtSSU rDNA, and a phylogenetic study of Botryosphaeria and associated anamorphic fungi. Mycol. Res. 2001, 105, 1033–1044. [Google Scholar] [CrossRef]
- Bárcenas-Peña, A.; Leavitt, S.D.; Grewe, F.; Lumbsch, H.T. Diversity of Xanthoparmelia (Parmeliaceae) species in Mexican xerophytic scrub vegetation, evidenced by molecular, morphological and chemistry data. An. Del Jardín Botánico De Madr. 2021, 78, e107. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES science gateway: A community resource for phylogenetic analyses. In Proceedings of the 2011 TeraGrid Conference: Extreme Digital Discovery TG ’11, Salt Lake City, UT, USA, 18–21 July 2011; Association for Computing Machinery: New York, NY, USA, 2011; pp. 1–8. [Google Scholar]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.-T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef]
- Gaiji, S.; Chavan, V.; Ariño, A.H.; Otegui, J.; Hobern, D.; Sood, R.; Robles, E. Content assessment of the primary biodiversity data published through GBIF network: Status, challenges and potentials. Biodivers. Inform. 2013, 8, 94–172. [Google Scholar]
- Linder, H.P.; de Klerk, H.M.; Born, J.; Burgess, N.D.; Fjeldså, J.; Rahbek, C. The partitioning of Africa: Statistically defined biogeographical regions in sub-Saharan Africa. J. Biogeogr. 2012, 39, 1189–1205. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Blázquez, M.; Hernández-Moreno, L.S.; Gasulla, F.; Pérez-Vargas, I.; Pérez-Ortega, S. The Role of Photobionts as Drivers of Diversification in an Island Radiation of Lichen-Forming Fungi. Front. Microbiol. 2022, 12, 4037. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Van Etten, J.; Cheng, J.; Mattiuzzi, M.; Greenberg, J.A. Package ‘Raster’; R package 3.6.14; R Foundation for Statistical Computing: Vienna, Austria, 2015; p. 734. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Gunderson, A.R.; Mahler, D.L.; Leal, M. Thermal niche evolution acroos replicated Anolis lizard adaptative radiations. Proc. R. Soc. B 2018, 285, 20172241. [Google Scholar] [CrossRef] [PubMed]
- Del Arco Aguilar, M.J.; González González, R.; Garzón Machado, V.; Pizarro Hernández, B. Actual and potential natural vegetation on the Canary Islands and its conservation status. Biodivers. Conserv. 2010, 19, 3089–3140. [Google Scholar] [CrossRef]
- Elix, J.A. Progress in the generic delimitation of Parmelia s. lat. lichens (Ascomycotina: Parmeliaceae) and a synoptic key to the Parmeliaceae. Bryologist 1993, 96, 359–383. [Google Scholar] [CrossRef]
- Elix, J.A. New species of Xanthoparmelia (lichenized Ascomycotina, Parmeliaceae) from Africa. Lichenologist 2002, 34, 283–291. [Google Scholar] [CrossRef]
- Elix, J.A.; Schumm, F. New species and new records in the lichen family Parmeliaceae (Ascomycota) from Macaronesia. Mycotaxon 2003, 86, 383–388. [Google Scholar]
- Sánchez-Pinto, L.; Rodríguez, S. Lichenes. In Lista Preliminar de Especies Silvestres de Cabo Verde (Hongos, Plantas y Animales Terrestres); Arechavaleta, M., Zurita, N., Marrero, M.C., Martín, J.L., Eds.; Consejería de Medio Ambiente y Ordenación Territorial, Gobierno de Canarias: La Laguna, Spain, 2005; pp. 27–33. [Google Scholar]
- Pérez-Vargas, I.; Hernández Padrón, C.; Elix, J.A. A new species of Xanthoparmelia (Ascomycota) from the Canary Islands. Lichenologist 2007, 38, 445–449. [Google Scholar] [CrossRef]
- Hernández Padrón, C.E.; Pérez-Vargas Lichenes, I. Lichenicolous Fungi. In Lista de Especies Silvestres de Canarias. Hongos, Plantas y Animales Terrestres; Arechavaleta, M., Rodríguez, S., Zurita, N., García, A., Coord., Eds.; Consejería de Medio Ambiente y Ordenación Territorial; Gobierno de Canarias: Santa Cruz de Tenerife, Spain, 2009; ISBN 978-84-89729-21-6. [Google Scholar]
- Bárcenas-Peña, A.; Leavitt, S.D.; Huang, J.P.; Grewe, F. Phylogenetic study and taxonomic revision of the Xanthoparmelia mexicana group, including the description of a new species (Parmeliaceae, Ascomycota). MycoKeys 2018, 40, 13–28. [Google Scholar] [CrossRef] [PubMed]
- Vančurová, L.; Peksa, O.; Němcová, Y.; Škaloud, P. Vulcanochloris (Trebouxiales, Trebouxiophyceae), a new genus of lichen photobiont from La Palma, Canary Islands, Spain. Phytotaxa 2015, 219, 118–132. [Google Scholar] [CrossRef]
- Barreno, E.; Muggia, L.; Chiva, S.; Molins, A.; Bordenave, C.; García-Breijo, F.; Moya, P. Trebouxia lynnae sp. nov. (former Trebouxia sp. TR9): Biology and biogeography of an epitome lichen symbiotic microalga. Biology 2022, 11, 1196. [Google Scholar] [CrossRef] [PubMed]
- Leliaert, F.; Smith, D.R.; Moreau, H.; Herron, M.D.; Verbruggen, H.; Delwiche, C.F.; De Clerck, O. Phylogeny and molecular evolution of the green algae. Crit. Rev. Plant Sci. 2012, 31, 1–46. [Google Scholar] [CrossRef]
- Škaloud, P.; Rindi, F.; Boedeker, C.; Leliaert, F. Freshwater Flora of Central Europe, Vol 13: Chlorophyta: Ulvophyceae (Süßwasserflora von Mitteleuropa, Bd. 13: Chlorophyta: Ulvophyceae); Springer: Berlin/Heidelberg, Germany, 2018; Volume 13. [Google Scholar]
- Muggia, L.; Nelsen, M.P.; Kirika, P.M.; Barreno, E.; Beck, A.; Lindgren, H.; Lumbsch, H.T.; Leavitt, S.D. Formally described species woefully underrepresent phylogenetic diversity in the common lichen photobiont genus Trebouxia(Trebouxiophyceae, Chlorophyta): An impetus for developing an integrated taxonomy. Mol. Phylogenet. Evol. 2020, 149, 106821. [Google Scholar] [CrossRef] [PubMed]
- Thüs, H.; Muggia, L.; Pérez-Ortega, S.; Favero-Longo, S.E.; Joneson, S.; O’Brien, H.; Nelsen, M.P.; Duque-Thüs, R.; Grube, M.; Friedl, T.; et al. Revisiting photobiont diversity in the lichen family Verrucariaceae (Ascomycota). Eur. J. Phycol. 2011, 46, 399–415. [Google Scholar] [CrossRef]
- Barber, J.C.; Francisco-Ortega, J.; Santos-Guerra, A.; Turner, K.G.; Jansen, R.K. Origin of Macaronesian Sideritis L. (Lamioideae: Lamiaceae) inferred from nuclear and chloroplast sequence datasets. Mol. Phylogenet. Evol. 2002, 23, 293–306. [Google Scholar] [CrossRef]
- Carine, M.; Russell, S.J.; Santos-Guerra, A.; Francisco-Ortega, J. Relationships of the Macaronesian and Mediterranean floras: Molecular evidence for multiple colonizations into Macaronesia and back-colonization of the continent in Convolvulus (Convolvulaceae). Am. J. Bot. 2004, 91, 1070–1085. [Google Scholar] [CrossRef] [PubMed]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation, 2nd ed.; Oxford University Press: Oxford, UK, 2007; ISBN 978-0-19-856611-3. [Google Scholar]
- Cronk, Q.C.B. Relict floras of Atlantic islands: Patterns assessed. Biol. J. Linn. Soc. 1992, 46, 91–103. [Google Scholar] [CrossRef]
- Olmstead, R.G.; Palmer, J.D. Implications for the Phylogeny, Classification, and Biogeography of Solanum from cpDNA Restriction Site Variation. Syst. Bot. 1997, 22, 19–29. [Google Scholar] [CrossRef]
- Mies, B.A. On the comparison of the flora and vegetation of the island groups of Socotra and Macaronesia. Bol. Mus. Mun. Funchal 1995, 4, 445–471. [Google Scholar]
- Kornhall, P.; Heidari, N.; Bremer, B. Selagineae and Manuleeae, two tribes or one? Phylogenetic studies in the Scrophulariaceae. Plant Syst. Evol. 2001, 228, 199–218. [Google Scholar] [CrossRef]
- Bohs, L.; Olmstead, R.G. A reassessment of Normania and Triguera (Solanaceae). Plant Syst. Evol. 2001, 228, 33–48. [Google Scholar] [CrossRef]
- Hjertson, M. Revision of the disjunct genus Campylanthus (Scrophulariaceae). Edinb. J. Bot. 2003, 60, 131–174. [Google Scholar] [CrossRef]
- Thiv, M.L.; Thulin, M.; Hjertson, M.; Kropf, M.; Linder, H.P. Evidence for a vicariant origin of Mafaronesian-Eritreo/Arabian disjunctions in Campylanthus Roth (Plantaginaceae). Mol. Phylogenet. Evol. 2010, 54, 607–616. [Google Scholar] [CrossRef]
- Pokorny, L.; Riina, R.; Mairal, M.; Meseguer, A.; Culshaw, V.; Cendoya, J.; Serrano, M.; Carbajal, R.; Ortiz, S.; Heuertz, M.; et al. Living on the edge: Timing of Rand Flora disjuncttions congruent with ongoing aridification in Africa. Front. Genet. 2015, 6, 1154. [Google Scholar] [CrossRef] [PubMed]
- Mairal, M.; Sanmartín, I.; Pellissier, L. Lineage-specific climatic niche drives the tempo of vicariance in the Rand Flora. J. Biogeogr. 2017, 44, 911–923. [Google Scholar] [CrossRef]
- Mairal, M.; Pokorny, L.; Aldasoro, J.J.; Alarcón, M.; Sanmartín, I. Ancient vicariance and climate-driven extinction explain continental disjunctions in Africa: The case of the Rand Flora genus Canarina (Campanulaceae). Mol. Ecol. 2005, 24, 1335–1354. [Google Scholar] [CrossRef] [PubMed]
- Mort, M.E.; Soltis, D.E.; Solsits, P.S.; Francisco-Ortega, J.; Santos-Guerra, A. Phylogenetics and Evolution of the Macaronesian Clade of Crassulaceae Inferred from Nuclear and Chloroplast Sequence Data. Syst. Bot. 2002, 27, 271–288. [Google Scholar]
- de Queiroz, A. The resurrection of oceanic dispersal in historical biogeography. Trends Ecol. Evol. 2005, 20, 68–73. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, F.; Guzmán, B.; Valido, A.; Vargas, P.; Arroyo, J. Late neogene history of the laurel tree (Laurus L.; Lauraceae) based on phylogeographical analyses of Mediterranean and Macaronesian populations. J. Biogeogr. 2009, 36, 1270–1281. [Google Scholar] [CrossRef]
- Navarro-Pérez, M.; Vargas, P.; Fernández-Mazuecos, M.; López, J.; Valtueña, F.J.; Ortega-Olivencia, A. Multiple windows of colonization to Macaronesia by the dispersal-unspecialized Scrophylaria since the Late Miocene. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 263–273. [Google Scholar] [CrossRef]
- de Nicolás, J.P.; Fernández-Palacios, J.M.; Ferrer, F.J.; Nieto, E. Inter-island floristic similarities in the Macaronesian Region. Vegetatio 1989, 84, 117–125. [Google Scholar] [CrossRef]
- Printzen, C.; Ekman, S.; Tonsberg, T. Phylogeography of Cavernularia hultenii: Evidence of slow genetic drift in a widely disjunct lichen. Mol. Ecol. 2003, 12, 1473–1486. [Google Scholar] [CrossRef]
- Buschbom, J. Migration between continents: Geographical structure and long-distance gene flow in Porpidia flavicunda (lichen-forming Ascomycota). Mol. Ecol. 2007, 16, 1835–1846. [Google Scholar] [CrossRef]
- Wirtz, N.; Printzen, C.; Lumbsch, H.T. The delimitation of Antarctic and bipolar species of neuropogonoid Usnea (Ascomycota, Lecanorales): A cohesion approach of species recognition for the Usnea perpusilla complex. Mycol. Res. 2008, 112, 472–484. [Google Scholar] [CrossRef]
- Geml, J.; Kauff, F.; Brochmann, C.; Lutzoni, F.; Laursen, G.A.; Redhead, S.A.; Taylor, D.L. Frequent circumarctic and rare transequatorial dispersals in the lichenised agaric genus Lichenomphalia (Hygrophoraceae, Basidiomycota). Fungal Biol. 2012, 116, 388–400. [Google Scholar] [CrossRef]
- Leavitt, S.D.; Fernandez-Mendoza, F.; Perez-Ortega, S.; Sohrabi, M.; Divakar, P.K.; Vondrak, J.; Lumbsch, H.T.; St Clair, L.L. Local representation of global diversity in a cosmopolitan lichen-forming fungal species complex (Rhizoplaca, Ascomycota). J. Biogeogr. 2013, 40, 1792–1806. [Google Scholar] [CrossRef]
- Bendiksby, M.; Mazzoni, S.; Jørgensen, M.H.; Halvorsen, R.; Holien, H. Combining genetic analyses of archived specimens with distribution modelling to explain the anomalous distribution of the rare lichen Staurolemma omphalarioides: Long-distance dispersal or vicariance? J. Biogeogr. 2014, 41, 2020–2031. [Google Scholar] [CrossRef]
- de Paz, G.A.; Cubas, P.; Crespo, A.; Elix, J.A.; Lumbsch, H.T. Transoceanic dispersal and subsequent diversification on separate continents shaped diversity of the Xanthoparmelia pulla group (Ascomycota). PLoS ONE 2012, 7, e39683. [Google Scholar] [CrossRef] [PubMed]
- Öckinger, E.; Niklasson, M.; Nilson, S.G. Is local distribution of the epiphytic lichen Lobatia pulmonaria limited by dispersal capacity or habitat quality? Biodivers. Conserv. 2005, 14, 759–773. [Google Scholar] [CrossRef]
- Ronnås, C.; Werth, S.; Ovaskainen, O.; Varkonyi, G.; Scheidegger, C.; Snäll, T. Discovery of long-distance gamete dispersal in a lichen-forming ascomycete. New Phytol. 2017, 216, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Tehler, A. The genera Dirina and Roccellina (Roccellaceae). Opera Bot. 1980, 70, 1–86. [Google Scholar] [CrossRef]
- Printzen, C.; Lumbsch, H.T. Molecular evidence for the diversification of extant lichens in the Late Cretaceous and Tertiary. Mol. Phylogenetics Evol. 2000, 17, 379–387. [Google Scholar] [CrossRef]
- Lücking, R.; Papong, K.; Thammathaworn, A.; Boonpragob, K. Historical biogeography and phenotype- phylogeny of Chroodiscus (lichenized Ascomycota: Ostropales: Graphidaceae). J. Biogeogr. 2008, 35, 2311–2327. [Google Scholar] [CrossRef]
- Schuster, M.; Duringer, P.; Ghienne, J.F.; Vignaud, P.; Mackaye, H.T.; Likius, A.; Brunet, M. The age of the Sahara Desert. Science 2006, 331, 821. [Google Scholar] [CrossRef] [PubMed]
- Pausata, F.S.; Gaetani, M.; Messori, G.; Berg, A.; de Souza, D.M.; Sage, R.F.; DeMenocal, P.B. The greening of the Sahara: Past changes and future implications. One Earth 2020, 2, 235–250. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
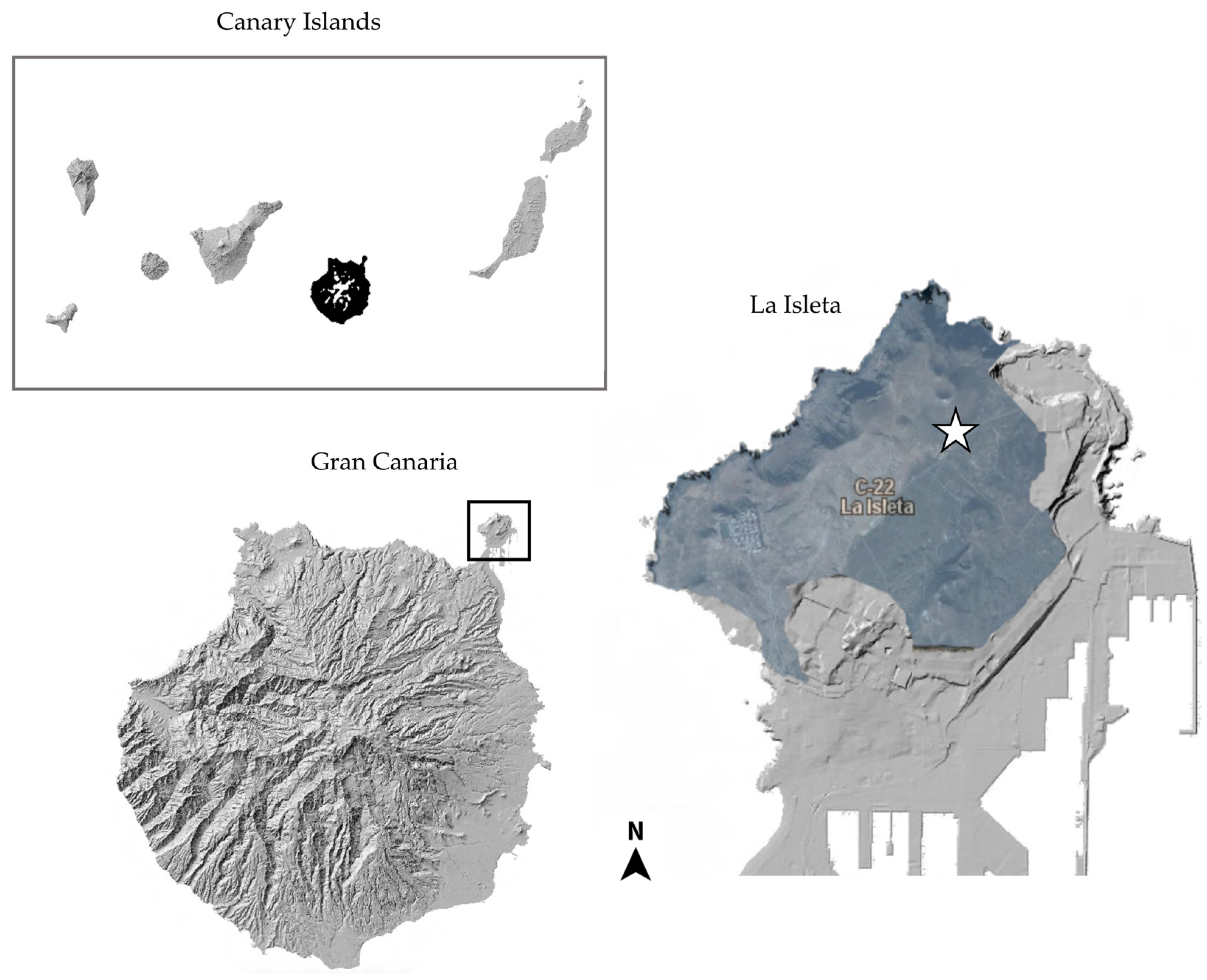{kind=link}
| Species | Geographic Origin | Voucher | Source | ITS | nuLSU | mtSSU |
|---|---|---|---|---|---|---|
| Xanthoparmelia hueana | Namibia | Baule 46511 | Leavitt et al. (2018) [34] | AY581090 | AY578956 | AY582326 |
| X. aff. krogiae | Kenya | Kirika 4637 | Leavitt et al. (2018) [34] | MG695502 | - | MG695750 |
| X. aff. krogiae | Kenya | MAF-Lich 17169 | Leavitt et al. (2018) [34] | JQ912361 | JQ912456 | - |
| X. patagonica | Argentina | Lumbsch 11036 | Leavitt et al. (2018) [34] | DQ980021 | DQ923670 | DQ923643 |
| X. phaeophana | Kenya | Kirika 4607 | Leavitt et al. (2018) [34] | - | MG695663 | MG695817 |
| X. aff. subramigera KE1 | Kenya | Kirika & Mugambi 3553 | Leavitt et al. (2018) [34] | MG695510 | MG695610 | MG695758 |
| X. aff. subramigera KE2 | Kenya | Kirika & Lumbsch 4117 | Leavitt et al. (2018) [34] | MG695514 | MG695615 | MG695763 |
| X. aff. subramigera KE3 | Kenya | Kirika & Lumbsch 4748 | Leavitt et al. (2018) [34] | MG695516 | - | MG695765 |
| X. aff. subramigera KE3 | Kenya | Kirika 3232 | Leavitt et al. (2018) [34] | MG695517 | MG695617 | MG695766 |
| X. aff. subramigera KE4 | Kenya | Kirika 3235 | Leavitt et al. (2018) [34] | MG695520 | MG695620 | MG695769 |
| X. aff. subramigera KE4 | Kenya | Kirika 3235 | Leavitt et al. (2018) [34] | MG695519 | MG695619 | MG695768 |
| X. aff. subramigera KE4 | Kenya | Kirika, Malombe & Matheka 3680 | Leavitt et al. (2018) [34] | MG695518 | MG695618 | MG695767 |
| X. aff. subramigera KE5 | Kenya | Kirika & Lumbsch 3811 | Leavitt et al. (2018) [34] | MG695521 | MG695621 | MG695770 |
| X. aff. subramigera KE6 | Kenya | Kirika & Lumbsch 4875 | Leavitt et al. (2018) [34] | MG695523 | MG695623 | MG695771 |
| X. aff. subramigera KE6 | Kenya | Kirika 2779 | Leavitt et al. (2018) [34] | MG695524 | MG695625 | MG695773 |
| X. aff. subramigera KE6 | Kenya | Kirika 4583 | Leavitt et al. (2018) [34] | MG695525 | MG695626 | MG695774 |
| X. aff. subramigera KE6 | Kenya | Kirika 4645 | Leavitt et al. (2018) [34] | MG695522 | MG695622 | - |
| X. aff. subramigera KE7 | Kenya | Kirika & Lumbsch 4812 | Leavitt et al. (2018) [34] | - | MG695628 | MG695776 |
| X. ramosae sp. nov. | Canary Islands | Pérez-Vargas LQ6 | This study | OR957381 | OR957383 | OR957385 |
| X. ramosae sp. nov. | Canary Islands | Pérez-Vargas LQ7 | This study | OR957382 | OR957384 | OR957386 |
| X. subramigera MX | Mexico | Ruiz-Cazares 1616 | Barcenas-Peña et al. (2021) [49] | MW553779 | - | - |
| X. subramigera MX | Mexico | Ruiz-Cazares 1619 | Barcenas-Peña et al. (2021) [49] | MW553780 | - | - |
| X. subramigera MX | Mexico | Ruiz-Cazares 1620 | Barcenas-Peña et al. (2021) [49] | MW553781 | - | - |
| X. subruginosa | Kenya | Kirika 3819 | Leavitt et al. (2018) [34] | MG695533 | MG695635 | MG695783 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Vargas, I.; Tuero-Septién, J.; Rancel-Rodríguez, N.M.; Pérez, J.A.; Blázquez, M. Patterns of Endemism in Lichens: Another Paradigm-Shifting Example in the Lichen Genus Xanthoparmelia from Macaronesia. J. Fungi 2024, 10, 166. https://doi.org/10.3390/jof10030166
Pérez-Vargas I, Tuero-Septién J, Rancel-Rodríguez NM, Pérez JA, Blázquez M. Patterns of Endemism in Lichens: Another Paradigm-Shifting Example in the Lichen Genus Xanthoparmelia from Macaronesia. Journal of Fungi. 2024; 10(3):166. https://doi.org/10.3390/jof10030166
Chicago/Turabian StylePérez-Vargas, Israel, Javier Tuero-Septién, Nereida M. Rancel-Rodríguez, José Antonio Pérez, and Miguel Blázquez. 2024. "Patterns of Endemism in Lichens: Another Paradigm-Shifting Example in the Lichen Genus Xanthoparmelia from Macaronesia" Journal of Fungi 10, no. 3: 166. https://doi.org/10.3390/jof10030166