
Genetic Diversity of Kazakhstani Equus caballus (Linnaeus, 1758) Horse Breeds Inferred from Microsatellite Markers
 , , and
, , and
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Laboratory Protocol
2.3. Population Genetic Structure
3. Results
Microsatellite Genotyping, and Population Genetic Diversity and Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lister, A.M.; Kadwell, M.; Kaagan, L.M.; Jordan, W.C.; Richards, M.B.; Stanley, H.F. Ancient and modern DNA in a study of horse domestication. Anc. Biomol. 1998, 2, 267–280. [Google Scholar]
- Vilà, C.; Leonard, J.A.; Götherström, A.; Marklund, S.; Sandberg, K.; Liden, K.; Wayne, R.K.; Ellegren, H. Widespread Origins of Domestic Horse Lineages. Science 2001, 291, 474–477. [Google Scholar] [CrossRef] [PubMed]
- Achilli, A.; Olivieri, A.; Soares, P.; Lancioni, H.; Hooshiar Kashani, B.; Perego, U.A.; Nergadze, S.G.; Carossa, V.; Santagostino, M.; Capomaccio, S.; et al. Mitochondrial genomes from modern horses reveal the major haplogroups that underwent domestication. Proc. Natl. Acad. Sci. USA 2012, 109, 2449–2454. [Google Scholar] [CrossRef]
- Warmuth, V.; Eriksson, A.; Bower, M.A.; Barker, G.; Barrett, E.; Hanks, B.K.; Li, S.; Lomitashvili, D.; Ochir-Goryaeva, M.; Sizonov, G.V.; et al. Reconstructing the origin and spread of horse domestication in the Eurasian steppe. Proc. Natl. Acad. Sci. USA 2012, 109, 8202–8206. [Google Scholar] [CrossRef]
- The Second Report on the State of the World’s Animal Genetic Resources for Food and Agriculture in Brief. Available online: https://www.fao.org (accessed on 23 May 2022).
- Horse Milk Has More Health Benefits Than Cow’s Milk—Researchers. Available online: https://guardian.ng/news/horse-milk (accessed on 12 March 2023).
- Horse Breeding in Kazakhstan: Development Prospects and Today’s Realities. Available online: https://world-nan.kz/blogs/innovatsionnyyi-granulyator-kormov-predstavili-na-eurotier-2022 (accessed on 10 March 2022). (In Russian).
- Dmitriez, N.; Ernst, L. KUSHUM (Kushumskaya), 330–331. In Animal Genetics Resources of the USSR; Dmitriez, N., Ernst, L., Eds.; Food and Agriculture Organization of the United Nations: Roma, Italy, 1989. [Google Scholar]
- Kushum Horse. Available online: https://world-animal.com/6333949-kushum-horse (accessed on 15 March 2022).
- Rzabayev, S.S. Mugalzhar Horse Breed; LLP Information and Printing Center—Kokzhiyek: Aktobe, Kazakhstan, 2007; p. 154. [Google Scholar]
- Satybaldin, A.A. Current state of horse breeding and horse sports in Kazakhstan. In Proceedings of the First International Conference, Kostanay, Kazakhstan, 2002. [Google Scholar]
- Moridi, M.; Masoudi, A.A.; Vaez Torshizi, R.; Hill, E.W. Mitochondrial DNA D-loop sequence variation in maternal lineages of Iranian native horses. Anim. Genet. 2013, 44, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Bowling, A.T.; Del Valle, A.; Bowling, M. A pedigree-based study of mitochondrial D-loop DNA sequence variation among Arabian horses. Anim. Genet. 2000, 31, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Csizmár, N.; Mihók, S.; Jávor, A.; Kusza, S. Genetic analysis of the Hungarian draft horse population using partial mitochondrial DNA D-loop sequencing. PeerJ 2018, 6, e4198. [Google Scholar] [CrossRef]
- Effa, K.; Rosenbom, S.; Han, J.; Dessie, T.; Beja-Pereira, A. Genetic Diversities and Historical Dynamics of Native Ethiopian Horse Populations (Equus caballus) Inferred from Mitochondrial DNA Polymorphisms. Genes 2021, 12, 155. [Google Scholar] [CrossRef]
- Almarzook, S.; Reissmann, M.; Brockmann, G.A. Diversity of mitochondrial DNA in three Arabian horse strains. J. Appl. Genet. 2017, 58, 273–276. [Google Scholar] [CrossRef]
- Hong, J.H.; Oh, C.H.; Kim, S.; Kang, I.U.; Shin, D.H. Genetic analysis of mitochondrial DNA from ancient Equus caballus bones found at archaeological site of Joseon Dynasty period capital area. Anim. Biosci. 2022, 35, 1141–1150. [Google Scholar] [CrossRef]
- McGahern, A.M.; Edwards, C.J.; Bower, M.A.; Heffernan, A.; Park, S.D.E.; Brophy, P.O.; Hill, E.W. Mitochondrial DNA sequence diversity in extant Irish horse populations and in ancient horses. Anim. Genet. 2006, 37, 498–502. [Google Scholar] [CrossRef] [PubMed]
- Yatkın, S.; Özdil, F.; Özkan Ünal, E.; Genç, S.; Kaplan, S.; Gürcan, E.K.; Arat, S.; Soysal, M.İ. Genetic Characterization of Native Donkey (Equus asinus) Populations of Turkey Using Microsatellite Markers. Animals 2020, 10, 1093. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Yun, S.W.; Cho, G.J. Assessment of genetic diversity using microsatellite markers to compare donkeys (Equus asinus) with horses (Equus caballus). Anim. Biosci. 2021, 34, 1460–1465. [Google Scholar] [CrossRef] [PubMed]
- Tozaki, T.; Kakoi, H.; Mashima, S.; Hirota, K.-I.; Hasegawa, T.; Ishida, N.; Miura, N.; Choi-Miura, N.-H.; Tomita, M. Population study and validation of paternity testing for Thoroughbred Horses by 15 microsatellite loci. J. Vet. Med. Sci. 2001, 63, 1191–1197. [Google Scholar] [CrossRef] [PubMed]
- Grilz-Seger, G.; Druml, T.; Neuditschko, M.; Dobretsberger, M.; Horna, M.; Brem, G. High-resolution population structure and runs of homozygosity reveal the genetic architecture of complex traits in the Lipizzan horse. BMC Genom. 2019, 20, 174. [Google Scholar] [CrossRef]
- Zhang, C.; Ni, P.; Ahmad, H.I.; Gemingguli, M.; Baizilaitibei, A.; Gulibaheti, D.; Fang, Y.; Wang, H.; Asif, A.R.; Xiao, C.; et al. Detecting the Population Structure and Scanning for Signatures of Selection in Horses (Equus caballus) From Whole-Genome Sequencing Data. Evol. Bioinform. 2018, 14, 1176934318775106. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Wang, S.; Zeng, G.; Guo, J.; Guo, M.; Dong, X.; Hua, G.; Liu, Y.; Wang, M.; Ling, Y.; et al. The Origin of a Coastal Indigenous Horse Breed in China Revealed by Genome-Wide SNP Data. Genes 2019, 10, 241. [Google Scholar] [CrossRef]
- Kargayeva, M.T.; Baimukhanov, D.A.; Nurbayev, S.D.; Baimukhanov, A.D.; Alikhanov, O.; Yusupbayev, Z. Identification of kazakh horses by microsatellite DNA using modern analytical methods. Bull. Natl. Acad. Sci. Repub. Kazakhstan 2020, 4, 55–61. [Google Scholar] [CrossRef]
- Gemingguli, M.; Iskhan, K.R.; Li, Y.; Qi, W.; Wunirifu, W.; Ding, L.Y.; Wumaierjiang, A. Genetic diversity and population structure of Kazakh horses (Equus caballus) inferred from mtDNA sequences. Genet. Mol. Res. 2016, 15, 15048618. [Google Scholar] [CrossRef]
- Seleuova, L.A.; Naimanov, D.K.; Jarowski, Z.; Aubakirov, M.Z.; Mustafin, B.M.; Safronova, O.S.; Baktybayev, G.T.; Turabaev, A.T.; Domatski, V.N. Population genetic characteristic of horses of Mugalzhar breed by STR-markers. Biomed. Res. 2018, 29, 3508–3511. [Google Scholar]
- Nguyen, T.B.; Paul, R.C.; Okuda, Y.; Le, T.N.A.; Pham, P.T.K.; Kaissar, K.J.; Kazhmurat, A.; Bibigul, S.; Bakhtin, M.; Kazymbet, P.; et al. Genetic characterization of Kushum horses in Kazakhstan based on haplotypes of mtDNA and Y chromosome, and genes associated with important traits of the horses. J. Equine. Sci. 2020, 31, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Kalashnikov, V.; Koveshnikov, V.; Sergienko, S. Prospects for the development of horse breeding in Russia in the new millennium. In Problems and Prospects of Horse Breeding in Russia at the Turn of the Century; Moscow, Russia, 2000; pp. 5–7. [Google Scholar]
- Canon, J.; Checa, M.L.; Carleos, C.; Vega-Pla, J.L.; Vallejo, M.; Dunner, S. The genetic structure of Spanish Celtic horse breeds inferred from microsatellite data. Anim. Genet. 2000, 31, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Bjornstad, G.; Gunby, E.; Roed, K.H. Genetic structure of Norwegian horse breeds. J. Anim. Breed. Genet. 2000, 117, 307–317. [Google Scholar] [CrossRef]
- Aberle, K.S.; Hamann, H.; Drogemuller, C.; Distl, O. Genetic diversity in German drought horse breeds compared with a group of primitive, riding and wild horses by means of microsatellite DNA markers. Anim. Genet. 2004, 35, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Zabek, T.; Fornal, A. Evaluation of the 17-plex STR kit for parentage testing of Polish coldblood and Hucul horses. Ann. Anim. Sci. 2009, 9, 363–372. [Google Scholar]
- Dimsoski, P. Development of a 17-plex microsatellite polymerase chain reaction kit for genotyping horses. Croat. Med. J. 2003, 44, 332–335. [Google Scholar]
- Marshal, T.C.; Slate, J.; Kruuk, L.E.B.; Pemberton, J.M. Statistical confidence for likelihood-based paternity inference in natural populations. Mol. Ecol. 1998, 7, 639–655. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2003, 16, 1099–1106. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Dieringer, D.; Schlötterer, C. Microsatellite analyser (MSA): A platform independent analysis tool for large microsatellite data sets. Mol. Ecol. Notes 2003, 3, 167–169. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Belkhir, K. GENETIX 4.05, Logiciel Sous Windows TM Pour la Génétique Des Populations. 2004. Available online: http://www.genetix.univ-montp2.fr/genetix/genetix.htm. (accessed on 17 December 2022).
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the numbers of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Jakobsson, M.; Rosenberg Noah, A. CLUMPP: A cluster matching and permutation program for dealing with label switching and multimodality in analysis of population structure. Bioinformatics 2007, 23, 1801–1806. [Google Scholar] [CrossRef]
- Rosenberg, N.A. Distruct: A program for the graphical display of population structure. Mol. Ecol. Notes 2004, 4, 137–138. [Google Scholar] [CrossRef]
- Yordanov, G.; Mehandjyiski, I.; Palova, N.; Atsenova, N.; Boyko, N.; Radoslavov, G.; Hristov, P. Genetic diversity and structure of the main Danubian horse paternal genealogical lineages based on microsatellite genotyping. Vet. Sci. 2022, 9, 333. [Google Scholar] [CrossRef]
- Jung, J.S.; Seong, J.; Lee, G.H.; Kim, Y.; An, J.H.; Yun, J.H.; Kong, H.S. Genetic diversity and relationship of Halla horse based on polymorphism in microsatellites. J. Anim. Reprod. Biotechnol. 2021, 36, 76–81. [Google Scholar] [CrossRef]
- Wright, S. The genetical structure of populations. Ann. Eugen. 1951, 15, 323–354. [Google Scholar] [CrossRef]
- Benhamadi, M.E.A.; Berber, N.; Benyarou, M.; Ameur, A.A.; Haddam, H.Y.; Piro, M.; Gaouar, S.B.S. Molecular Characterization of Eight Horse Breeds in Algeria Using Microsatellite Markers. Biodiversitas 2020, 21, d210923. [Google Scholar] [CrossRef]
- Dorji, J.; Tamang, S.; Tshewang, T.; Dorji, T.; Dorji, T.Y. Genetic diversity and population structure of three traditional horse breeds of Bhutan based on 29 DNA microsatellite markers. PLoS ONE 2018, 13, e199376. [Google Scholar] [CrossRef] [PubMed]
- Yüceer, B.; Erdoğan, M.; Yaralı, C.; Özarslan, B.; Özbeyaz, C. Genetic diversity between Rahvan (pacing) horses in Turkey. Ankara Üniv. Vet. Fak. Derg. 2016, 63, 201–210, (In Turkısh). [Google Scholar]
- Ling, Y.H.; Ma, Y.H.; Guan, W.J.; Cheng, Y.J.; Wang, Y.P.; Han, J.L.; Mang, L.; Zhao, Q.J.; He, X.H.; Pu, Y.B.; et al. Evaluation of the genetic diversity and population structure of Chinese indigenous horse breeds using 27 microsatellite markers. Anim. Genet. 2011, 42, 56–65. [Google Scholar] [CrossRef]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef]
- Behl, R.; Behl, J.; Gupta, N.; Gupta, S.C. Genetic relationships of five Indian horse breeds using a microsatellite markers. Animal 2007, 1, 483–488. [Google Scholar] [CrossRef]
- Solis, A.; Jugo, B.M.; Meriaux, J.C.; Iriondo, M.; Mazon, L.I.; Aguirre, A.I.; Vicario, A.; Estomba, A. Genetic diversity within and among four south European Native Horse breeds based on microsatellite DNA analysis: Implications for conservation. J. Hered. 2005, 96, 670–678. [Google Scholar] [CrossRef]
- Voronkova, V.N.; Nikolaeva, E.A.; Piskunov, A.K.; Babayan, O.V.; Takasu, M.; Tozaki, T.; Svishcheva, G.R.; Stolpovsky, Y.A. Assessment of Genetic Diversity and Structure of Russian and Mongolian Autochthonous Horse Breeds Using Nuclear and Mitochondrial DNA Markers. Russ. J. Genet. 2022, 58, 927–943. [Google Scholar] [CrossRef]
- Blokhina, N.V.; Khrabrova, L.A.; Gavrilicheva, I.S. Application of modern technologies in identifying distinctive features in the subpopulation of Novoaltaiskaya hoses. IOP Conf. Ser. Earth Environ. Sci. 2021, 624, 012019. [Google Scholar] [CrossRef]
- Zaitcev, A.M.; Gavrilicheva, I.S.; Blohina, N.V.; Khrabrova, L.A.; Kokorina, N.B. Assessment of the population structure of horses of the Priobskaya breed based on modern technologies. International Conference on World Technological trends in Agribusiness. IOP Conf. Ser. Earth Environ. Sci. 2021, 624, 012032. [Google Scholar] [CrossRef]
- Iwańczyk, E.; Juras, R.; Cholewiński, G.; Cothran, E.G. Genetic structure and phylogenetic relationships of the Polish Heavy horse. J. Appl Genet. 2006, 47, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Juras, R.; Cothran, E.G.; Klimas, R. Genetic Analysis of Three Lithuanian Native Horse Breeds. Acta. Agric. Scand. 2003, 53–54, 180–185. [Google Scholar] [CrossRef]
- Funk, S.M.; Guedaoura, S.; Juras, R.; Raziq, A.; Landolsi, F.; Luís, C.; Martínez, A.M.; Mayaki, A.M.; Mujica, F.; Oom, M.D.M.; et al. Major inconsistencies of inferred population genetic structure estimated in a large set of domestic horse breeds using microsatellites. Ecol. Evol. 2020, 10, 4261–4279. [Google Scholar] [CrossRef] [PubMed]
- Wright, S. Evolution and the genetics of populations, vol. 4. In Variability Within and among Natural Populations; University of Chicago Press: Chicago, IL, USA, 1978; p. 580. [Google Scholar]
- Petersen, J.L.; Mickelson, J.R.; Cothran, E.G.; Andersson, L.S.; Axelsson, J.; Bailey, E.; Bannasch, D.; Binns, M.M.; Borges, A.S.; Brama, P.; et al. Genetic diversity in the modern horse illustrated from genome-wide SNP data. PLoS ONE 2013, 8, e54997. [Google Scholar] [CrossRef] [PubMed]
- Marchiori, C.M.; Pereira, G.L.; Maiorano, A.M.; Rogatto, G.M.; Assoni, A.D.; Silva, J.A.V., II; Chardulo, L.A.L.; Curi, R.A. Linkage disequilibrium and population structure characterization in the cutting and racing lines of Quarter Horses bred in Brazil. Livest. Sci. 2019, 219, 45–51. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
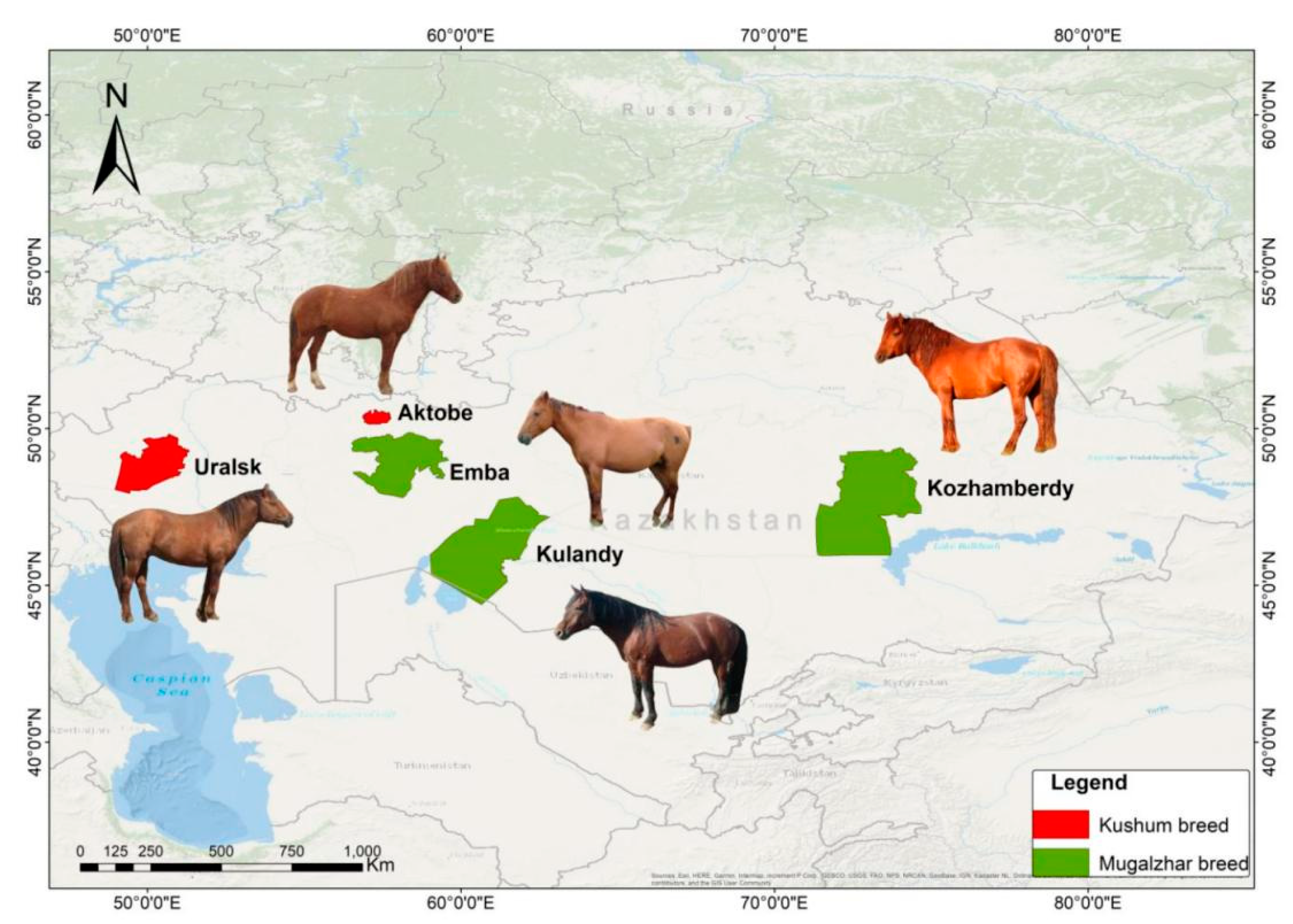
| N | Population | Ns | Location |
|---|---|---|---|
| Kushum breed | |||
| 1 | Uralsk type | 76 | Zhangali district, W KZ |
| 2 | Aktobe type | 74 | Aktobe region, W KZ |
| Mugalzhar breed | |||
| 6 | Emba type | 93 | Aktobe region, W KZ |
| 7 | Kulandy type | 101 | Kyzylorda region, SW KZ |
| Kozhamberdy type | |||
| 3 | Maupas interbreed line | 29 | Karaganda region, C KZ |
| 4 | Meiman interbreed line | 28 | Karaganda region, C KZ |
| 5 | Mesker interbreed line | 34 | Karaganda region, C KZ |
| Locus | NA | NE | I | HO | HE | UHE | FIS | FIT | FST | PIC |
|---|---|---|---|---|---|---|---|---|---|---|
| VHL20 | 10.57 | 6.026 | 1.991 | 0.855 | 0.830 | 0.839 | −0.031 | 0.004 | 0.034 | 0.813 |
| HTG4 | 6.423 | 2.900 | 1.338 | 0.634 | 0.642 | 0.650 | 0.013 | 0.044 | 0.032 | 0.629 |
| AHT4 | 8.429 | 5.701 | 1.867 | 0.746 | 0.823 | 0.832 | 0.093 | 0.125 | 0.035 | 0.822 |
| HMS7 | 6.571 | 3.911 | 1.496 | 0.667 | 0.728 | 0.736 | 0.083 | 0.112 | 0.031 | 0.734 |
| AHT5 | 7.000 | 4.908 | 1.695 | 0.779 | 0.796 | 0.805 | 0.021 | 0.051 | 0.030 | 0.784 |
| HMS6 | 7.571 | 4.660 | 1.717 | 0.652 | 0.783 | 0.793 | 0.168 | 0.206 | 0.046 | 0.788 |
| ASB23 | 9.857 | 5.678 | 1.918 | 0.502 | 0.822 | 0.837 | 0.389 | 0.415 | 0.042 | 0.713 |
| ASB2 | 10.00 | 6.334 | 1.927 | 0.842 | 0.806 | 0.896 | −0.045 | 0.033 | 0.074 | 0.627 |
| HTG7 | 7.000 | 3.568 | 1.453 | 0.751 | 0.712 | 0.720 | −0.054 | −0.010 | 0.041 | 0.686 |
| HMS3 | 8.143 | 4.676 | 1.711 | 0.566 | 0.784 | 0.793 | 0.278 | 0.306 | 0.040 | 0.786 |
| LEX3 | 9.571 | 5.728 | 1.912 | 0.479 | 0.814 | 0.825 | 0.422 | 0.439 | 0.047 | 0.768 |
| Mean | 8.286 | 4.917 | 1.730 | 0.679 | 0.776 | 0.793 | 0.121 | 0.157 | 0.041 | 0.741 |
| Source of Variation | d.f. | SS | MS | Est. Var. | % of Variation |
|---|---|---|---|---|---|
| Among populations | 6 | 240.411 | 40.069 | 0.290 | 6% |
| Among individuals | 428 | 2270.183 | 5.304 | 1.074 | 24% |
| Within individuals | 435 | 1372.500 | 3.155 | 3.155 | 70% |
| Total | 869 | 3883.094 | 4.520 | 100% | |
| FST = 0.064 (p ≥ 0.001); FIS = 0.254 (p ≥ 0.001); FIT = 0.302 (p ≥ 0.001) | |||||
| Uralsk | Aktobe | Maupas | Meiman | Mesker | Emba | Kulandy | |
|---|---|---|---|---|---|---|---|
| Uralsk | |||||||
| Aktobe | 0.045 | ||||||
| Maupas | 0.037 | 0.021 | |||||
| Meiman | 0.035 | 0.020 | 0.008 | ||||
| Mesker | 0.041 | 0.021 | 0.010 | 0.014 | |||
| Emba | 0.033 | 0.014 | 0.014 | 0.014 | 0.021 | ||
| Kulandy | 0.047 | 0.025 | 0.022 | 0.021 | 0.028 | 0.021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orazymbetova, Z.; Ualiyeva, D.; Dossybayev, K.; Torekhanov, A.; Sydykov, D.; Mussayeva, A.; Baktybayev, G. Genetic Diversity of Kazakhstani Equus caballus (Linnaeus, 1758) Horse Breeds Inferred from Microsatellite Markers. Vet. Sci. 2023, 10, 598. https://doi.org/10.3390/vetsci10100598
Orazymbetova Z, Ualiyeva D, Dossybayev K, Torekhanov A, Sydykov D, Mussayeva A, Baktybayev G. Genetic Diversity of Kazakhstani Equus caballus (Linnaeus, 1758) Horse Breeds Inferred from Microsatellite Markers. Veterinary Sciences. 2023; 10(10):598. https://doi.org/10.3390/vetsci10100598
Chicago/Turabian StyleOrazymbetova, Zarina, Daniya Ualiyeva, Kairat Dossybayev, Aibyn Torekhanov, Dauren Sydykov, Aizhan Mussayeva, and Gabiden Baktybayev. 2023. "Genetic Diversity of Kazakhstani Equus caballus (Linnaeus, 1758) Horse Breeds Inferred from Microsatellite Markers" Veterinary Sciences 10, no. 10: 598. https://doi.org/10.3390/vetsci10100598