
A Flow Cytometry-Based Assessment of the Genomic Size and Ploidy Level of Wild Musa Species in India
by

Rithesh B. Natarajan
1,
Pooja Pathania
2,
Hardeep Singh
2,
Anuradha Agrawal
1,2,3 and
Rajkumar Subramani
1,2,* 1
Division of Plant Genetic Resources, ICAR-Indian Agricultural Research Institute, Pusa Campus, New Delhi 110012, India
2
ICAR-National Bureau of Plant Genetic Resources, Pusa Campus, New Delhi 110012, India
3
Indian Council of Agricultural Research, Pusa Campus, New Delhi 110012, India
*
Author to whom correspondence should be addressed.
Plants 2023, 12(20), 3605; https://doi.org/10.3390/plants12203605
Submission received: 4 August 2023
/
Revised: 12 October 2023
/
Accepted: 15 October 2023
/
Published: 18 October 2023
Abstract
:The genome size variation is an important attribute in evolutionary and species characterization. Musa L. is regarded as one of the taxonomically complicated genera within the order Zingiberales, with more than 75 species from wild seeded to seedless cultivars that may be diploid, triploid or tetraploid. The knowledge of total nuclear DNA content in terms of genome size and ploidy level in wild species of Musa is absolutely important in evolutionary and genomic studies. Methods: In this paper, chromosome spreading was performed via protoplast isolation and a fast air-dry dropping method and flow cytometry were used with Raphanus sativus L. (Brassicaceae) as a standard for ploidy and genome size estimation. Results: The results showed that genome size (2C) varied amongst Musa species, based on the ratio of G1 peak positions. The lowest genome size (2C) was found in M. balbisiana var. andamanica (1.051 ± 0.060 pg) and the highest genome size (2C) was recorded for Musa ABB.cv. Meitei-hei (1.812 ± 0.108 pg) for the section Eumusa. Among the species belonging to the section Rhodochlamys, M. rosae had the lowest 2C content of 1.194 ± 0.033 pg whereas the highest nuclear DNA content (2C) was observed in M. velutina (1.488 ± 0.203 pg). Cytogenetic analysis revealed that the chromosome number of 14 wild Musa species was 2n = 22, while 1 species—Ensete glaucum—showed a chromosome number of 2n = 18 (diploid), and for 3 species, the chromosome number was 2n = 33 (triploids). An association study based on the Pearson correlation coefficient showed 2C nuclear DNA content was significant and positively correlated with ploidy level (R = 0.9) and chromosome number (R = 0.84). Conclusions: The present study provides reliable information on the genome size and ploidy level of wild Musa species from the Indian region through flow cytometric analysis, which could be further utilized in taxonomic and crop improvement programs. For the first time, the nuclear DNA content of eight wild diploid and three triploid Indian species were estimated and reported. Genome size could be an effective indicator in identification of species and evolutionary studies in Musa with varying ploidy levels and morphological similarities.
1. Introduction
Banana (Musa spp.) represents one of the world’s major staple fruit crops and is an important export commodity for millions of people living in tropical and subtropical regions [1]. It is a significant export commodity for various developing nations and plays a crucial role in their national trade, contributing to their socioeconomic development. In 2021, the annual global banana production was reported to be around 124.97 million tons (mt), out of which the highest amount of 33.06 mt was produced by India (https://www.fao.org/faostat/en/#data, accessed on 8 May 2013). Bananas are monocotyledonous plants belonging to the family Musaceae under the order Zingiberales with three genera: Musa L., Ensete Horan., and Musella (Franch.) H.W. Li. [2] The largest genus, Musa, comprises about 75 species and is mainly distributed in Southeast Asian region [1,3]. Ensete, a smaller genus, is present in both Asia and Africa [4,5], whereas the monotypic genus Musella is native to Southwest China [6]. The genus Musa was divided into five sections—Australimusa, Callimusa, Eumusa, Incertae, and Rhodochlamys—based on their chromosome number and morphological characters [4,7]. This traditional classification of Musa has been reappraised based on numerous molecular studies merging Eumusa and Rhodochlamys into the section Musa and Australimusa and Callimusa into the section Callimusa [3,8]. The majority of the edible banana cultivars have originated from inter- and intra-specific hybridization of two wild diploid species (2n = 2x = 22), namely M. acuminata Colla (A genome) and M. balbisiana Colla (B genome). Other species, such as M. schizocarpa N.W. Simmonds (S genome) and M. textilis Nee (T genome), also contributed to the origin of some edible banana clones to some extent [9,10]. The combinations of these genomes have resulted in various genotypes of cultivated edible banana clones, exhibiting different genomic composition and ploidy levels such as AA, AB, AAA, AAB, ABB, AAAB, AABB, and ABBB [11]. In India, the genus is represented by 37 wild taxa—of which 20 are endemic—and is largely distributed in northeastern states and followed by Western Ghats, Eastern Ghats, and Andaman and Nicobar Islands [12,13]. However, the family Musaceae is well known for its vast genetic diversity and thought to have originated from south and Southeast Asia [13,14]. Wild seeded bananas play a vital role in enhancing banana crop improvement, serving as a valuable repository of a diverse gene pool that harbors resistant genes against significant diseases, pests, and abiotic stresses. Despite being a major fruit crop, only M. acuminata and M. balbisiana have been used for breeding novel varieties [15,16]. Recently, a search for traits in evolutionarily related Musa species revealed a socioeconomic importance of wild bananas in banana breeding programs. However, to our knowledge, only a few wild species have been studied and to a limited extent, so little is known about their nuclear genomes.
The use of molecular tools provides an efficient method for determination of genome composition and has been extensively applied in taxonomic classification of Musa species. One of the basic characteristics of the taxonomic studies of higher plants is their nuclear genome size, which serves as an essential feature in classification and provides useful insights about various species [17]. The genome size or DNA C value is essential and remains a key component in the genomic studies, phylogeny, and species classification [18,19]. Considering genome size in conjunction with chromosome numbers and ploidy status can offer valuable perspectives on the evolutionary relationships between closely related taxa in wild plants [20]. Genome size can have significant variation within the same genus and among different individuals within a species [19,21]. The nuclear genome size (equal to 1C) can be expressed either in picograms (pg) or megabase pairs (1 pg = 978 Mbp), corresponding to the method of estimation [22]. So far, genome size in Musa spp. has been estimated to be relatively small (1C, ~ 600 Mbp) and divided between 11 chromosomes [23]. These estimations were focused only on M. acuminata, M. balbisiana, triploid, and tetraploid cultivars. Consequently, there is an urgent necessity to extend the knowledge of nuclear genome sizes to the other Musa species as well. Although different methods have been used to estimate genome sizes in plants, flow cytometry (FC) serves as a primary method for the estimation of nuclear DNA content (2C) and ploidy. In comparison to the traditional chromosome-counting technique, FC holds an advantage as it allows for the screening of numerous plants in a short timeframe [23,24]. It has also become the method of choice due to the ease of sample preparation, relatively low cost, and the ability to analyze high numbers of nuclei; it can also be applied to any plant tissue [25]. FC determines the relative DNA content of each nucleus by quantifying the fluorescence emitted by each stained nucleus; the estimation also requires a species with known genome size as an internal or external standard [24]. Furthermore, the genome size also plays a significant role in distinguishing hidden Musa species where the variation in ploidy is known. Therefore, this study was conducted to determine the nuclear DNA content, ploidy level, and chromosome count of wild Musa in India with the aim to investigate the genome size variation using FC analysis.
2. Results
2.1. Chromosome Number Analysis
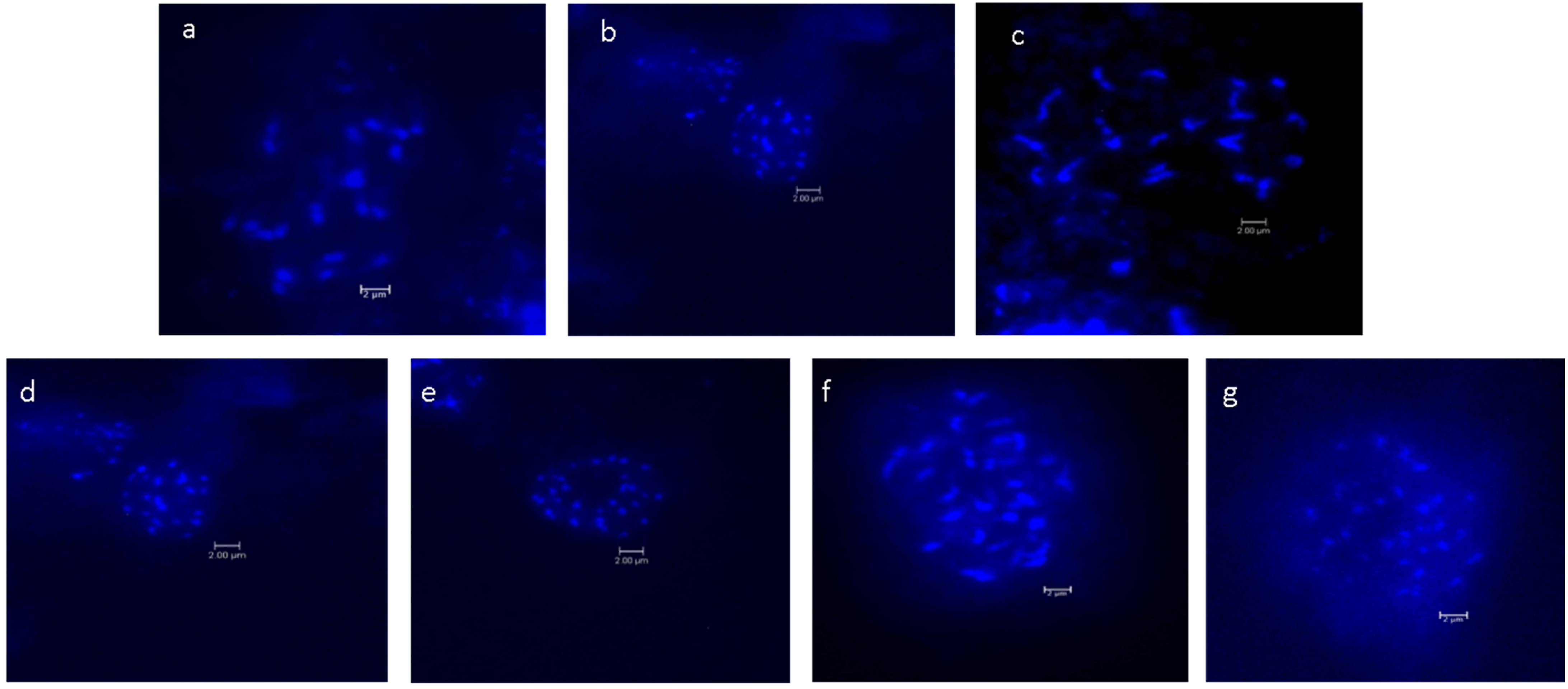
The chromosome numbers of the Musa species and cultivars are presented in Table 1. Images of metaphase spread reported for the first time for seven Musa species and cultivars are shown in Figure 1. The chromosome count revealed that 14 wild Musa species were confirmed to be diploids, comprising the sections Eumusa and Rhodochlamys with chromosome number 2n = 22. In the section Eumusa, 3 Musa species were triploids with a chromosome number 2n = 33. In the genus Ensete, the species Ensete glaucum showed a different chromosome number, 2n = 18.
2.2. Ploidy Level Analysis
This study showed that out of 18 accessions, 14 Musa accessions were diploids (2n = 2x = 22) and 3 were triploids (2n = 3x = 33). One species of genus Ensete, Ensete glaucum, was diploid (2n = 2x = 18) (Table 1). A significant difference was observed in fluorescent intensity corresponding to different ploidy levels; for each value of ploidy, the fluorescent intensity of the G1 peak differed significantly between diploids and triploids. However, the study also showed that most of the wild Musa accessions in both sections (Eumusa and Rhodochlamys) were diploid, but three cultivars in the section Eumusa were triploids.
2.3. Variation in Genome Size Values
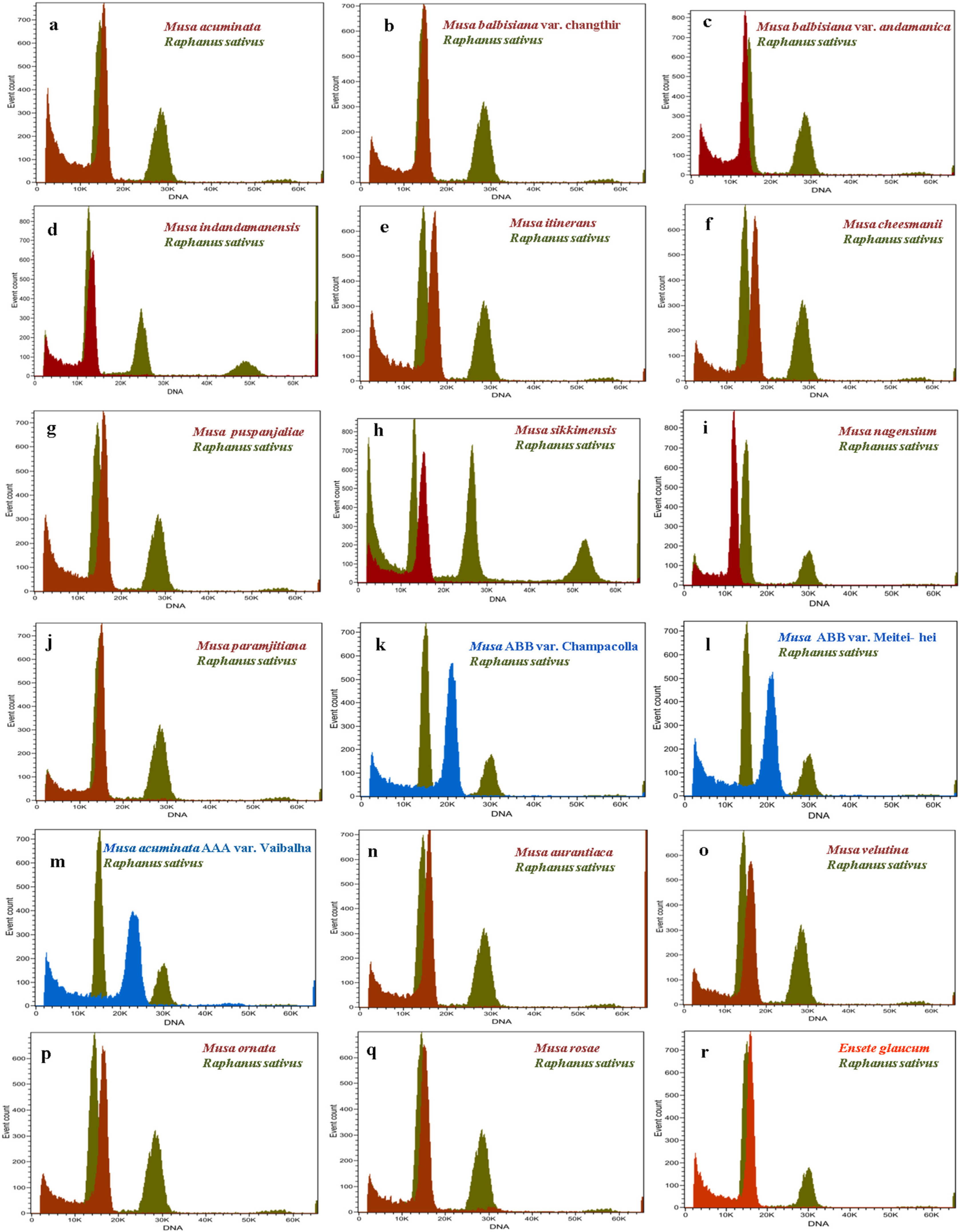
The amount of nuclear DNA was estimated after flow cytometric analysis of propidium iodide-stained nuclei, and the means 2C values corresponding to Musa species are presented in Table 1. The histograms of relative nuclear DNA content with two dominant peaks corresponding to G1 nuclei of Musa sp. and Raphanus sativus are shown in Figure 2. The coefficient of variation (CV) for G1 of all Musa sample peaks was less than five, indicating the good quality of sample preparation and reliability of the results. The analysis revealed the nuclear DNA content as 2C values based on the ratio of G1 peak positions, which ranged from 1.051 to 1.812 pg for the section Eumusa. The lowest nuclear DNA size was observed in the diploid taxon M. balbisiana var. andamanica with 1.051 pg and the highest nuclear DNA content was recorded for the triploid Musa ABB var. Meitei-hei with 1.812 pg in the Eumusa section. The section Rhodochlamys showed a similar range of nuclear DNA content, ranging from 1.194 to 1.488 pg. The lowest nuclear DNA content was recorded for M. rosae (1.194 pg) and the highest for M. velutina (1.488 pg). Ensete glaucum, an outgroup in the section, had 2C nuclear DNA content of 1.349 pg. The FC analysis of 2C nuclear DNA contents of two sections, Eumusa and Rhodochlamys, showed overlapping results with little difference. The three triploid Musa accessions—namely, Musa AAB. var. Champacolla, Musa ABB. var. Meitei-hei, and Musa acuminata AAA. Vaibalhla—showed a higher range of nuclear DNA content of 1.802, 1.812, and 1.741 pg, respectively, than diploid species from both sections. The mean 2C values of Raphanus sativus and wild Musa species from the Eumusa and Rhodochlamys sections were close together, and a merged inflorescence peak was observed when Raphanus sativus was used as a reference standard. Considerable variation in nuclear DNA content was also observed among Musa accessions representing both sections.
The study showed the calculated average genome size (1C content) of all Musa accession, which ranged from 514.1 to 885.9 Mbp. The Musa balbisiana var. andamanica showed the lowest 1C content of 514.1 Mbp among all the accessions, while the Musa ABB. var. Meitei-hei showed the highest 1C content with 885.9 Mbp. Based on the Pearson correlation coefficient, the 2C nuclear DNA content showed a strong significance and positively correlated with ploidy level (R = 0.9) (Figure 3A) and chromosome number (R = 0.84) (Figure 3B) among the Musa species.
3. Discussion
Southeast Asia is considered one of the world’s biodiversity hotspots for Musa spp. India has a significant amount of banana genetic diversity in the wild and is known as the center of origin for interspecific hybrids worldwide. The basic genomic information of wild Musa species originated from India would be of much importance for future banana improvement programs. We have determined the chromosome number, ploidy level, and genome size of wild Musa species in India.
3.1. Chromosome Number and Ploidy Status
The number of somatic chromosomes of Musa will be important when designating and confirming the species ploidy level. The hard and rigid cell walls from the root system of cultured Musa species were a limiting factor for preparing chromosome spread [26] through the traditional squash method, so protoplast isolation and fast air-dry dropping chromosome preparation was performed in this study. Cytogenetic analysis confirmed the chromosome number of wild Musa species from sections Eumusa and Rhodochlamys as diploids (2n = 2x = 22), and triploid cultivars were found to be 2n = 3x = 33, ensuring the use of basic chromosome numbers for classifying the Musa species [4]. The genus Ensete and its species with monocarpic habits are sister genera close to Musa in the family Musaceae, but with the basic chromosome number x = 9 [4,27]. The Ensete glaucum is a diploid with a chromosome number of 2n = 2x = 18. Although chromosome counting is accurate and applied primarily to plants with large chromosomes, challenges remain with regard to the banana’s chromosome; due to its small, indistinguishable size and high level of metaphase condensation, it is hard to obtain a high quality of chromosome spread [27,28]. There is limited information concerning the structural characteristics of chromosomes and karyotypes for the Musa species, and detailed information about the chromosomes’ number and their size, shape, and arrangement within this genus remains scarce [29]. A variety of indirect methods for determining banana ploidy level have been reported [30,31]. However, this method depends on statistical analysis and is not accurate. In the present study, the flow cytometry method is further applied to confirm the ploidy status of Musa spp.
FC provides a rapid and efficient method for large-scale determination of ploidy level in Musaceae [32]. Christelová et al. [33] used the flow cytometry method to rapidly estimate and genotype nearly half of the ITC accessions (495 accessions) of the global Musa germplasm collection. In the present study, the ploidy level was determined by measuring the peak position of sample nuclei in comparison with the peak position of reference nuclei, supplemented with DAPI during flow cytometry analysis. The fluorescence intensity was significantly different for each Musa species, in which the ploidy level increased with increased fluorescent intensity. We also observed a shift in the peak position to the right as an increase in the ploidy level of Musa species. Furthermore, the flow cytometry analysis for determining ploidy reconfirms via the chromosome counting method that the ploidy status of the 14 Musa and one Ensete species are diploids and three cultivars are triploids. Therefore, the knowledge on Musa ploidy is valuable for taxonomy and crop improvement as it plays an important role in classifying individual species.
3.2. Analysis of Genome Size
Nuclear genome size is considered a basic characteristic in classifying the species and providing valuable information on taxonomy and the evolutionary relationship of the higher plant [16,34]. Earlier reports on the flow cytometry technique also suggested that it could be applied to the analysis of the nuclear DNA content of unknown samples with respect to known reference standards and their ability to differentiate between closely related species, even of various ploidy levels [16,23]. The present results showed that the nuclear DNA content of Musa balbisiana (1.046 pg) was smaller than Musa acuminata (1.220 pg). Kamaté et al. [35] reported 1.16 pg for Musa balbisiana (BB genome), 1.27 pg for Musa acuminata (AA genome), and also recorded the difference at their subspecies level of approximately 11%. Our present study used genome size as a key component to determine the intra- and interspecific variation and the relationship between wild Musa species in India. Current analysis showed a wide range of nuclear DNA content in the sections Eumusa and Rhodochlamys. Several researchers noted a close relationship between species of the sections Eumusa and Rhodochlamys, both morphologically [27] and at molecular level [36]. Our results also showed a close relationship between two investigated sections. The high range of variation in the nuclear DNA content (2C/pg) can also enrich the Musa C- value database. Čížková et al. [36] studied the nuclear DNA content of 21 diploid accessions representing sections of the genus Musa and showed the 2C DNA content in the section Musa ranged from 1.217 to 1.315 pg. Investigations on the genome size of genera Musa and Ensete showed overlap among their genome sizes, with a range from 0.61 to 0.69 pg/1C in Eumusa and from 0.6 to 0.66 pg/1C in Rhodochlamys [16,17]. Similarly, our present study showed 2C nuclear DNA values in the Eumusa and Rhodochlamys sections exhibited a degree of overlap with little difference. The three triploid Musa accessions showed a higher range of nuclear DNA content than diploid species in both sections. Our study also showed the calculated average 1C value of all Musa species ranged from 514.1 Mbp to 885.9 Mbp in both the Eumusa and Rhodochlamys sections. Lysảk et al. [37] also observed variation in the genome size of the genus Musa. The FC method previously used to estimate the genome size resulted in a clear distinction between M. acuminata and M. balbisiana with genome size of ~0.6–0.64 pg/1C and ~0.55–0.57 pg/1C, respectively, and is closely related to our findings.
The positive correlation between chromosome number and genome size in the Indian wild Musa species is similar to that of other studies on cultivated and wild species of Musa [24]. The intraspecific variation in genome size is attributed to variation in the tandem repeats, transposable elements (TEs), polyploidization, and recombination rate [38,39]. Likewise, in Musa 55% of the genome is composed of various repetitive and non-coding sequences [40], while the nuclear genome in bananas contains coding genes every 6.4 to 6.9 kb [41,42]. The variation in repetitive sequence was found to be a major factor in differentiation in the genus Musa, even at subspecies level [43,44]. The difference in genome size between M. acuminata and M. balbisiana has been attributed to a variation in the copy number of repetitive DNA sequences such as “Monkey” [45] and Radka5 [46]. The systematic studies on the newly described endemic species of Musa from India revealed all of them belong to either the balbisiana or acuminata complex. Along with sequence data, the nuclear genome size of newly described Musa species will provide valuable information on genomic studies, evolutionary relationship, and species classification.
4. Materials and Methods
4.1. Plant Material
In vitro-rooted plantlets of 17 Musa and one Ensete species were obtained from Tissue Culture and Cryopreservation Unit, ICAR-National Bureau of Plant Genetic Resources (NBPGR), New Delhi (Table 2). Raphanus sativus cv Saxa (2C = 1.11 pg) reference standard was obtained from Institute of Experimental Botany, Czech Republic.
4.2. Cytogenetic Studies
Mitotic metaphase spreads were prepared according to the method described by Doleželová et al. [47], with minor modifications. Actively growing Musa root tips were pre-treated in 0.002 M of 8-hydroxyquinoline for 4 h at room temperature and then fixed for 24 h in 3:1 ethanol: acetic acid overnight and stored in 70% ethanol. Fixed roots were washed with 1X enzyme buffer solution (40 mL of 100 mM Citric Acid, 60 mL of 100 mM Trisodium Citrate and pH adjusted to 4.8, and diluted 1:10 in distilled water) and digested in the enzyme solution (containing 2% w/v cellulase, 3% v/v pectinase and 1X enzyme buffer) and kept in a bath-marie at 37° C for 2.5 h. Digested root tips were washed thrice with 1X enzyme buffer and cell suspension was prepared using 3:1 acetic acid: ethanol. The suspension was dropped onto a clean dry ice-cold microscopic slide placed on a hot plate (55 °C) from a height of 30 to 40 cm. The suspension was allowed to spread out and air dry [48]. Finally, the slides were examined under a fluorescence microscope (Leica DM6 B) and the images captured using the Leica application suite (LAS) X 3.8 software.
4.3. Flow Cytometry—Determination of Nuclear Genome Size and Ploidy Status
A small amount of leaf sample (~50 mg) was taken in a glass Petri dish and 500 μL of extraction buffer (CyStain PI absolute P kit, Sysmex, Germany) was added. The sample was finely chopped using a sharp scalpel blade for 30 s. and filtered through 20 μm CellTrics filter into a sample tube. For genome size estimation, the chopped samples were incubated in 2 mL of staining solution, supplemented with Propidium Iodide and Rnase (CyStain PI Absolute P kit, Sysmex, Germany), and incubated in the dark on ice for 30 min. The sample was analyzed in a flow cytometer (CyFlow Ploidy Analyser, Sysmex, Germany) under red channel (532 nm). Raphanus sativus was used as a reference standard. The nuclear genome size of samples was determined using the formula given by Doležel et al. [49]. 2C nuclear DNA content = Reference 2C value × Sample 2C mean peak position Reference 2C mean peak position.
The mean nuclear DNA content was then calculated for each plant. The genome size, which represents one copy of nuclear genetic information (equal to 1C), was further determined considering 1 pg = 0.978 × 109 bp [22].
1C (Mbp) = nuclear DNA content (pg) × (0.978 × 109)/2
To improve accuracy, the nuclear DNA content was determined for each Musa sample in three replicates, enabling the standard error to be calculated.
For ploidy determination, staining buffer supplemented with DAPI was added to the sample after the nuclei extraction step. The experiment was performed in three replicates. Relative fluorescence intensity of stained nuclei was analyzed using a flow cytometer (CyFlow Ploidy Analyser) under a UV channel. Standardization of flow cytometry was carried out using a confirmed diploid Musa species, Musa balbisiana (ITC-0565), obtained from the Musa Germplasm Transit Centre. Musa balbisiana was used as reference standard for ploidy screening to assess the ploidy status of other species under analysis. The ploidy of Musa species was determined by comparing the fluorescence values between the peak positions of samples to that of reference standards. The Musa samples were analyzed in the flow cytometry under UV channel and a minimum of 5000 particles were measured for both reference and samples.
Analyses of variance (ANOVA) were performed to analyze the variation in nuclear DNA size (R 4.2.1 software- Stat package and agricolae). The significance level = 0.001 was used. Pearson correlation analysis was performed to learn the associations among nuclear DNA content, chromosome number and ploidy status using R 4.2.1 software (Package-ggpubr and ggplot2).
5. Conclusions
Our study showed that the identification of genome size and ploidy level is important for taxonomy and crop improvement programs. There was no clear gap in nuclear DNA content between the sections Eunusa and Rhodochlamys. Furthermore, this study showed that nuclear DNA content in relation with the chromosome numbers and ploidy status provided valuable information on the evolutionary relationships and characterization of Musa species. We observed a significant difference in the nuclear genome size and ploidy status of wild Musa species belonging to the Eumusa and Rhodochlamys sections. Our study also showed the positive correlation between nuclear DNA content and ploidy level and emphasizes that flow cytometry can be used for rapid and precise estimation of ploidy as a metric for genome characterization.
Author Contributions
Conceptualization, R.S.; Methodology, R.B.N., P.P., H.S., A.A. and R.S.; Software, R.B.N. and P.P.; Validation, R.S.; Formal analysis, R.B.N. and P.P.; Supervision, R.S.; Resources, A.A.; Data curation, R.S.; Writing—original draft preparation, R.B.N. and P.P.; Writing—review and editing, R.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
All the data are available in the manuscript.
Acknowledgments
We are thankful to the Director, ICAR- National Bureau of Plant Genetic Resources for providing facilities to carry out this research work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Šimoníková, D.; Čížková, J.; Zoulová, V.; Christelová, P.; Hřibová, E. Advances in the Molecular Cytogenetics of Bananas, Family Musaceae. Plants 2022, 11, 482. [Google Scholar] [CrossRef] [PubMed]
- Kress, W.J. The phylogeny and classification of the Zingiberales. Ann. Missouri. Bot. Gard. 1990, 77, 698–721. [Google Scholar] [CrossRef]
- Häkkinen, M. Reappraisal of sectional taxonomy in Musa (Musaceae). Taxon 2013, 62, 809–813. [Google Scholar] [CrossRef]
- Cheesman, E.E. Classification of the bananas: The genus Ensete Horan and the genus Musa L. Kew Bull. 1947, 2, 97–117. [Google Scholar] [CrossRef]
- Baker, R.; Simmonds, N.W. The genus Ensete in Africa. Kew Bull. 1953, 8, 405–416. [Google Scholar] [CrossRef]
- Häkkinen, M.; Vare, H. Typification and check-list of Musa L. names (Musaceae) with nomenclatural notes. Adansonia 2008, 30, 63–112. [Google Scholar]
- Šimoníková, D.; Němečková, A.; Karafiátová, M.; Uwimana, B.; Swennen, R.; Doležel, J.; Hřibová, E. Chromosome painting facilitates anchoring reference genome sequence to chromosomes in situ and integrated karyotyping in banana (Musa spp.). Front. Plant Sci. 2019, 10, 1503. [Google Scholar] [CrossRef] [PubMed]
- Christelová, P.; Valárik, M.; Hřibová, E.; Van den Houwe, I.; Channelière, S.; Roux, N.; Doležel, J. A platform for efficient genotyping in Musa using microsatellite markers. AoB Plants 2011, 2011, plr024. [Google Scholar] [CrossRef]
- Čížková, J.; Hřibová, E.; Humplíková, L.; Christelová, P.; Suchánková, P.; Doležel, J. Molecular analysis and genomic organization of major DNA satellites in banana (Musa spp.). PLoS ONE 2013, 8, e54808. [Google Scholar] [CrossRef]
- Němečková, A.; Christelová, P.; Čížková, J.; Nyine, M.; van den Houwe, I.; Svačina, R.; Uwimana, B.; Swennen, R.; Doležel, J.; Hřibová, E. Molecular and cytogenetic study of East African Highland Banana. Front. Plant. Sci. 2018, 9, 1371. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, N.W.; Shepherd, K. The taxonomy and origins of the cultivated bananas. J. Linn. Soc. Bot. 1955, 55, 302–312. [Google Scholar] [CrossRef]
- Joe, A.; Sabu, M. Revision of Indian Musaceae; IAAT, Calicut University: Kerala, India, 2019; pp. 1–345. ISBN 81-901637-5-2. [Google Scholar]
- Uma, S.; Saraswathi, M.S.; Backiyarani, S.; Durai, P.; Agrawal, A. Unravelling the potential of Musa genetic resources for benefit of global banana community. In Banana & Plantains: Leading-Edge Research and Developments; Uma, S., Mayil Vaganan, M., Agrawal, A., Eds.; Volume 1: Germplasm Diversity, and Breeding; ICAR-National Research Centre for Banana: Tiruchirappalli, India, 2020; pp. 1–44. ISBN 978-81-945318-0-7. [Google Scholar]
- Prasad, K.; Joe, A.; Bheemalingappa, M.; Rao, B.R.P. Musa sabuana (Musaceae): A new species from Andaman and Nicobar Islands. Indian J. For. 2013, 36, 151–153. [Google Scholar] [CrossRef]
- Laliberte, B.; De Langhe, E. Chapter 6: Musa collections around the world. In Global Strategy for the Conservation and Use of Musa Genetic Resources; vol C-Management; Bioversity International: Rome, Italy, 2016; pp. 44–70. [Google Scholar]
- Bartoš, J.; Alkhimova, O.; Doleželová, M.; De Langhe, E.; Doležel, J. Nuclear genome size and genomic distribution of ribosomal DNA in Musa and Ensete (Musaceae): Taxonomic implications. Cytogenet. Genome Res. 2005, 109, 50–57. [Google Scholar] [CrossRef]
- Bennett, M.D.; Johnston, S.; Hodnett, G.L.; Price, H.J. Allium cepa L. cultivars from four continents compared by flow cytometry show nuclear DNA constancy. Ann. Bot. 2000, 85, 351–357. [Google Scholar] [CrossRef]
- Leitch, I.J.; Bennett, M.D. Genome size and its uses: The impact of flow cytometry. In Flow Cytometry with Plant Cells: Analysis of Genes, Chromosomes and Genomes; Wiley-VCH: Weinheim, Germany, 2007; pp. 153–176. [Google Scholar]
- Dai, S.-F.; Zhu, X.-G.; Hutang, G.-R.; Li, J.-Y.; Tian, J.-Q.; Jiang, X.-H.; Zhang, D.; Gao, L.-Z. Genome Size Variation and Evolution Driven by Transposable Elements in the Genus Oryza. Front. Plant Sci. 2022, 13, 921937. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.; Weiss-Schneeweiss, H.; Temsch, E.M.; So, S.; Myeong, H.H.; Jang, T.S. Genome size and chromosome number evolution in Korean Iris L. species (Iridaceae Juss.). Plants 2020, 9, 1284. [Google Scholar] [CrossRef] [PubMed]
- Tsutsui, N.D.; Suarez, A.V.; Spagna, J.C.; Johnston, J.S. The evolution of genome size in ants. BMC. Evol. Biol. 2008, 8, 64. [Google Scholar] [CrossRef] [PubMed]
- Doležel, J.; Bartoš, J.; Voglmayr, H.; Greilhuber, J. Nuclear DNA content and genome size of trout and human. Cytom. Part A 2003, 51, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Doležel, J.; Doleželová, M.; Novák, F.J. Flow cytometric estimation of nuclear DNA amount in diploid bananas (Musa acuminata and M. balbisiana). Biol. Plantarum. 1994, 36, 351–357. [Google Scholar] [CrossRef]
- Doležel, J.; Bartoš, J. Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef]
- Rampácková, E.; Mrázová, M.; Cížková, J.; Necas, T. Pomological Traits and Genome Size of Prunus armeniaca L. Considering to Geographical Origin. Horticulturae 2022, 8, 199. [Google Scholar] [CrossRef]
- Doležel, J.; Dolezelová, M.; Roux, N.; van Den Houwe, I. A novel method to prepare slides for high resolution chromosome studies in Musa spp. Infomusa 1998, 7, 3–4. [Google Scholar]
- Simmonds, N.W. Notes on banana taxonomy. Kew Bull. 1960, 14, 198–212. [Google Scholar] [CrossRef]
- Pillay, M.; Tenkouano, A. Genomes, Cytogenetics and Flow Cytometry of Musa. In Banana Breeding: Progress and Challenges; Pillay, M., Tenkouano, A., Eds.; CRC Press: Boca Raton, FL, USA, 2011. [Google Scholar]
- Dehery, S.K.; Panda, E.; Saha, P.R.; Sinha, R.K.; Das, A.B. Chromosome diversity and karyotype asymmetry analysis in four cultivated triploid and three diploid wild genotypes of Musa from North-East India. Nucleus 2020, 64, 167–179. [Google Scholar] [CrossRef]
- Vandenhout, H.; Ortiz, R.; Vuylsteke, D.; Swennen, R.; Bai, K.V. Effect of ploidy on stomatal and other quantitative traits in plantain and banana hybrids. Euphytica 1995, 83, 117–122. [Google Scholar] [CrossRef]
- Tenkouano, A.; Crouch, J.H.; Crouch, H.K.; Vuylsteke, D. Ploidy determination in Musa germplasm using pollen and chloroplast characteristics. HortScience 1998, 33, 889–890. [Google Scholar] [CrossRef]
- Doleželová, M.; Doležel, J.; van der Houwe, I.; Roux, N.; Swennen, R. Focus on the Musa collection: Ploidy levels revealed. Infomusa 2005, 14, 34–36. [Google Scholar]
- Christelová, P.; de Langhe, E.; Hřibová, E.; Čížková, J.; Sardos, J.; Hušáková, M.; van den houwe, I.; Sutanto, A.; Kepler, A.K.; Swennen, R.; et al. Molecular and cytological characterization of the global Musa germplasm collection provides insights into the treasure of banana diversity. Biodivers. Conserv. 2017, 26, 801–824. [Google Scholar] [CrossRef]
- Loureiro, J.; Trávníček, P.; Rauchova, J.; Urfus, T.; Vit, P.; Štech, M.; Castro, S.; Suda, J. The use of flow cytometry in the biosystematics, ecology and population biology of homoploid plants. Preslia 2010, 82, 3–21. [Google Scholar]
- Kamaté, K.; Brown, S.; Durand, P.; Bureau, J.M.; De Nay, D.; Trinh, T.H. Nuclear DNA content and base composition in 28 taxa of Musa. Genome 2001, 44, 622–627. [Google Scholar] [CrossRef]
- Čížková, J.; Hřibová, E.; Christelova, P.; Van den Houwe, I.; Häkkinen, M.; Roux, N.; Swennen, R.; Doležel, J. Molecular and cytogenetic characterization of wild Musa species. PLoS ONE 2015, 10, e0134096. [Google Scholar] [CrossRef]
- Lysák, M.A.; Doleželová, M.; Horry, J.P.; Swennen, R.; Doležel, J. Flow cytometric analysis of nuclear DNA content in Musa. Theor. Appl. Genet. 1999, 98, 1344–1350. [Google Scholar] [CrossRef]
- Tiley, G.P.; Burleigh, J.G. The relationship of recombination rate, genome structure, and patterns of molecular evolution across angiosperms. BMC Evol. Biol. 2015, 15, 194. [Google Scholar] [CrossRef]
- Bennetzen, J.L.; Ma, J.; Devos, K.M. Mechanisms of recent genome size variation in flowering plants. Ann. Bot. 2005, 95, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Hřibová, E.; Doleželová, M.; Town, C.D.; Macas, J.; Doležel, J. Isolation and characterization of the highly repeated fraction of the banana genome. Cytogenet. Genome Res. 2007, 119, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Aert, R.; Sági, L.; Volckaert, G. Gene content and density in banana (Musa acuminata) as revealed by genomic sequencing of BAC clones. Theor. Appl. Genet. 2004, 109, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Cheung, F.; Town, C.D. A BAC end view of the Musa acuminata genome. BMC Plant Biol. 2007, 7, 29. [Google Scholar] [CrossRef] [PubMed]
- Baurens, F.C.; Noyer, J.L.; Lanaud, C.; Lagoda, P.J.L. Use of competitive PCR to assay copy number of repetitive elements in banana. Mol. Gen. Genet. 1996, 253, 57–64. [Google Scholar] [CrossRef]
- Baurens, F.C.; Noyer, J.L.; Lanaud, C.; Lagoda, P.J.L. Assessment of a species-septic element (Brep 1) in banana. Theor. Appl. Genet. 1997, 95, 922–931. [Google Scholar] [CrossRef]
- Balint-Kurti, P.J.; Clendennen, S.K.; Doleželová, M.; Valárik, M.; Doležel, J.; Beetham, P.R.; May, G.D. Identication and chromosomal localization of the monkey retrotransposon in Musa sp. Mol. Gen. Genet. 2000, 263, 908–915. [Google Scholar] [CrossRef]
- Valárik, M.; Šimková, H.; Hřibová, E.; Šafář, J.; Doleželová, M.; Doležel, J. Isolation, characterization and chromosome localization of repetitive DNA sequences in bananas (Musa spp.). Chromosome Res. 2002, 10, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Doleželová, M.; Valárik, M.; Swennen, R.; Horry, J.P.; Doležel, J. Physical mapping of the 18S-25S and 5S ribosomal RNA genes in diploid bananas. Biol. Plantarum. 1998, 41, 497–505. [Google Scholar] [CrossRef]
- Aliyeva-Schnorr, L.; Ma, L.; Houben, A. A Fast Air-dry Dropping Chromosome Preparation Method Suitable for FISH in Plants. J. Vis. Exp. 2015, 16, e53470. [Google Scholar] [CrossRef]
- Doležel, J.; Greilhuber, J.; Suda, J. Estimation of nuclear DNA content in plants using flow cytometry. Nat. Protoc. 2007, 2, 2233–2244. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Mitotic metaphase chromosomes of Musa: (a). M. balbisiana var. andamanica, (b). M. indandamanensis, (c). M. sikkimensis, (d). M. paramjitiana, (e). M. AAB var. Champacolla, (f). M. ABB var. Meitei-hei, (g). M. AAA var. Vaibalha (scale bar 2 um).
Figure 1.
Mitotic metaphase chromosomes of Musa: (a). M. balbisiana var. andamanica, (b). M. indandamanensis, (c). M. sikkimensis, (d). M. paramjitiana, (e). M. AAB var. Champacolla, (f). M. ABB var. Meitei-hei, (g). M. AAA var. Vaibalha (scale bar 2 um).

Figure 2.
Histogram of relative nuclear DNA content, with the dominant peak corresponding to G1 nuclei of Musa and Raphanus sativus obtained after flow cytometry analysis. (a). M. acuminata, (b). M. balbisiana var. Changthir, (c). M. balbisiana var. andamanica, (d). M. indandamanensis, (e). M. itinerans, (f). M. cheesmanii, (g). M. puspanjaliae, (h). M. sikkimensis, (i). M. nagensium, (j). M. paramjitiana, (k). M. AAB cv. Champacolla, (l). M. ABB cv. Meitei-hei, (m). M. AAA var. Vaibalha, (n). M.aurantiaca, (o). M.velutina, (p). M.ornata, (q). M. rosae, and (r). Ensete glaucum.
Figure 2.
Histogram of relative nuclear DNA content, with the dominant peak corresponding to G1 nuclei of Musa and Raphanus sativus obtained after flow cytometry analysis. (a). M. acuminata, (b). M. balbisiana var. Changthir, (c). M. balbisiana var. andamanica, (d). M. indandamanensis, (e). M. itinerans, (f). M. cheesmanii, (g). M. puspanjaliae, (h). M. sikkimensis, (i). M. nagensium, (j). M. paramjitiana, (k). M. AAB cv. Champacolla, (l). M. ABB cv. Meitei-hei, (m). M. AAA var. Vaibalha, (n). M.aurantiaca, (o). M.velutina, (p). M.ornata, (q). M. rosae, and (r). Ensete glaucum.

Figure 3.
Relationship between nuclear 2C DNA content of the Musa species (A) with ploidy status, (B) with chromosome number.
Figure 3.
Relationship between nuclear 2C DNA content of the Musa species (A) with ploidy status, (B) with chromosome number.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Nuclear DNA content, genome size, ploidy level, and chromosome count of investigated accessions from Musa and Ensete genera and two standard species (Raphanus sativus and Musa balbisiana cultivar ITC0565).
Table 1.
Nuclear DNA content, genome size, ploidy level, and chromosome count of investigated accessions from Musa and Ensete genera and two standard species (Raphanus sativus and Musa balbisiana cultivar ITC0565).
| Section | Species | 2C Nuclear DNA (pg) Average ± SE | Genome Size 1C (Mbp) | Ploidy | Chromosome Number (2n) |
|---|---|---|---|---|---|
| Raphanus sativus | 1.11 ± 0.00 | 542.8 | 2 | 18 | |
| Eumusa | Musa acuminata | 1.220 ± 0.087 | 596.6 | 2 | 22 |
| Musa balbisiana (ITC0565) | 1.046 ± 0.034 | 511.4 | 2 | 22 | |
| Musa balbisiana var. Changthir | 1.150 ± 0.071 | 562.5 | 2 | 22 | |
| Musa balbisiana var. andamanica | 1.051 ± 0.060 | 514.1 | 2 | 22 | |
| Musa indandamanensis | 1.171 ± 0.080 | 572.7 | 2 | 22 | |
| Musa itinerans | 1.349 ± 0.031 | 659.7 | 2 | 22 | |
| Musa cheesmani | 1.373 ± 0.232 | 671.5 | 2 | 22 | |
| Musa puspanjaliae | 1.221 ± 0.046 | 596.9 | 2 | 22 | |
| Musa sikkimensis | 1.359 ± 0.072 | 664.6 | 2 | 22 | |
| Musa nagensium | 1.093 ± 0.016 | 534.3 | 2 | 22 | |
| Musa paramjitiana | 1.152 ± 0.045 | 563.3 | 2 | 22 | |
| Musa AAB var. Champacolla | 1.802 ± 0.063 | 881.2 | 3 | 33 | |
| Musa ABB var. Meitei-hei | 1.812 ± 0.108 | 885.9 | 3 | 33 | |
| Musa acuminata AAA var. Vaibalhla | 1.741 ± 0.109 | 851.3 | 3 | 33 | |
| Rhodochlamys | Musa aurantiaca | 1.238 ± 0.029 | 605.2 | 2 | 22 |
| Musa velutina | 1.488 ± 0.203 | 727.7 | 2 | 22 | |
| Musa ornata | 1.292 ± 0.027 | 631.6 | 2 | 22 | |
| Musa rosae | 1.194 ± 0.033 | 584.0 | 2 | 22 | |
| Ensete glaucum | 1.349 ± 0.093 | 659.8 | 2 | 18 |
Statistical analysis was performed using mean values calculated for individual plants (n = 3) and significance level α = 0.01.
Table 2.
List of Musa and Ensete species used in the present study.
| S.No | Species | Accession Detail | Genomic Constitution | Section | Place of Collection | Crop/Special Characteristics |
|---|---|---|---|---|---|---|
| 1 | Musa acuminata | IC633381 | AA | Eumusa | Mizoram | Seeded Banana |
| 2 | Musa balbisiana | ITC0565 | BB | Eumusa | Belgium | Ploidy reference standard |
| 3 | Musa balbisiana var. changthir | IC833382 | BB | Eumusa | Mizoram | Seeded Banana |
| 4 | Musa balbisiana var. andamanica | IC630992 | BB | Eumusa | Andaman and Nicobar Islands | Seeded Banana |
| 5 | Musa indandamanensis | IC631162 | - | Eumusa | Andaman and Nicobar Islands | Wild Banana with 11 mts height, sweet with orange fruit plup |
| 6 | Musa itinerans | - | - | Eumusa | - | Wild banana with Pink Fruit and spreading rhizomatous roots |
| 7 | Musa cheesmani | - | - | Eumusa | - | Wild banana with Black hard seed |
| 8 | Musa puspanjaliae | - | - | Eumusa | - | Largest seeded banana |
| 9 | Musa sikkimensis | - | - | Eumusa | Sikkim | Red striped leaves |
| 10 | Musa nagensium | IC627969 | - | Eumusa | Arunachal Pradesh | Wild Banana |
| 11 | Musa paramjitiana | IC628650 | - | Eumusa | Andaman and Nicobar Islands | Sweet and sour tasting fruit with boat shape and bulb shaped seeds |
| 12 | Musa AAB. var. Champacolla | - | AAB | Eumusa | - | Cultivar |
| 13 | Musa ABB. Meitei-hei | - | ABB | Eumusa | - | Cultivar |
| 14 | Musa acuminata AAA. Vaibalhla | - | AAA | Eumusa | - | Cultivar |
| 15 | Musa aurantiaca | IC627978 | - | Rhodochlamys | Nagaland | Dwarf Banana with bright orange inflorescence |
| 16 | Musa velutina | IC636539 | - | Rhodochlamys | Assam | Pink Fruiting Banana/Hairy Banana |
| 17 | Musa ornata | IC633379 | - | Rhodochlamys | Mizoram | Flowering Banana |
| 18 | Musa rosae | - | - | Rhodochlamys | - | Wild ornamental Banana |
| 19 | Ensete glaucum | IC633380 | - | Mizoram | Snow Banana |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Natarajan, R.B.; Pathania, P.; Singh, H.; Agrawal, A.; Subramani, R. A Flow Cytometry-Based Assessment of the Genomic Size and Ploidy Level of Wild Musa Species in India. Plants 2023, 12, 3605. https://doi.org/10.3390/plants12203605
AMA Style
Natarajan RB, Pathania P, Singh H, Agrawal A, Subramani R. A Flow Cytometry-Based Assessment of the Genomic Size and Ploidy Level of Wild Musa Species in India. Plants. 2023; 12(20):3605. https://doi.org/10.3390/plants12203605
Chicago/Turabian StyleNatarajan, Rithesh B., Pooja Pathania, Hardeep Singh, Anuradha Agrawal, and Rajkumar Subramani. 2023. "A Flow Cytometry-Based Assessment of the Genomic Size and Ploidy Level of Wild Musa Species in India" Plants 12, no. 20: 3605. https://doi.org/10.3390/plants12203605
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.