
Diversity and Ecology of Lobophora Species Associated with Coral Reef Systems in the Western Gulf of Thailand, including the Description of Two New Species
 , ,
, ,
Abstract
:1. Introduction
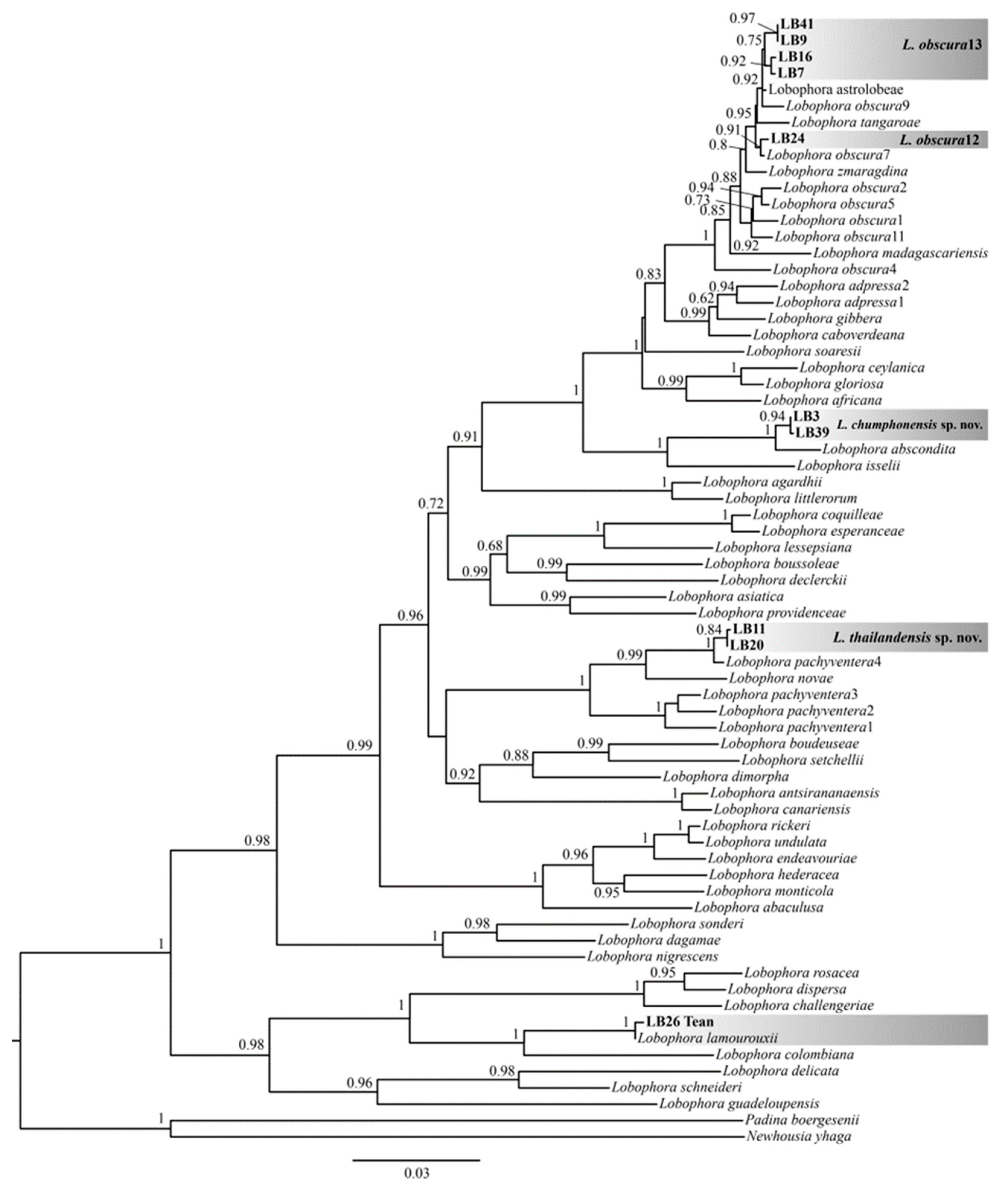
2. Results
2.1. Taxonomic Results
2.1.1. Lobophora chumphonensis sp. nov. A.M.Klomjit and C.W.Vieira (Figure 2g,h, Table 1)

2.1.2. Lobophora thailandensis sp. nov. A.M.Klomjit and C.W.Vieira (Figure 2i,j, Table 1)
2.2. Morphological, Anatomical, and ECOLOGICAL Characteristics

{kind=link}
{kind=link}
{kind=link}
| Characteristics | Algal Species | CV (%) | ||||
|---|---|---|---|---|---|---|
| L. chumphonensis sp. nov. | L. lamourouxii | L. obscura12 | L. obscura13 | L. thailandensis sp. nov. | ||
| Thickness | 36.57 | |||||
| Average | 90.3 ± 9.8 b | 95.1 ± 9.8 b | 190.8 ± 28.5 a | 177.0 ± 13.0 ab | 98.4 ± 9.1 b | |
| Min–Max | 75–105 | 86–113 | 162–240 | 156–191 | 87–110 | |
| Number of cells | 32.06 | |||||
| Average | 8 | 5 | 10 | 8 | 8 | |
| Min–Max | 5–11 | 5 | 8–12 | 5–11 | 4–12 | |
| Number of dorsal cells | 42.30 | |||||
| Average | 4 | 2 | 5 | 4 | 4 | |
| Min–Max | 2–6 | 2 | 4–6 | 2–6 | 1–7 | |
| Number of ventral cells | 33.33 | |||||
| Average | 3 | 2 | 4 | 3 | 3 | |
| Min–Max | 2–4 | 2 | 3–5 | 2–4 | 2–4 | |
| Medulla length | 15.22 | |||||
| Average | 78.9 ± 7.3 a | 81.3 ± 4.4 a | 65.7 ± 16.7 a | 71.1 ± 8.0 a | 71.7 ± 5.6 a | |
| Min–Max | 71–90 | 75–87 | 36–82.5 | 59–83 | 63–78 | |
| Medulla height | 41.68 | |||||
| Average | 21.3 ± 3.5 ab | 36 ± 4.14 a | 20.4 ± 2.9 ab | 13.5 ± 1.3 b | 13.8 ± 1.7 b | |
| Min–Max | 21–25.5 | 30–42 | 18–24 | 12–15 | 12–17 | |
| Medulla width | 27.58 | |||||
| Average | 27.9 ± 6.9 a | 17.4 ± 2.8 a | 26.7 ± 5.6 a | 27.9 ± 4.3 a | 19.8 ± 2.0 a | |
| Min–Max | 21–39 | 14–21 | 20–35 | 23–35 | 17–23 | |
| Dorsal height | 60.30 | |||||
| Average | 28.8 ± 4.3 abc | 22.2 ± 1.4 bc | 67.5 ± 6.7 a | 64.8 ± 7.0 ab | 12.6 ± 2.2 c | |
| Min–Max | 24–36 | 21–24 | 60–75 | 40.5–54 | 9–15 | |
| Ventral height | 41.75 | |||||
| Average | 24.3 ± 3.5 b | 21.0 ± 4.2 b | 50.4 ± 6.1 a | 36.9 ± 5.9 ab | 20.7 ± 3.6 b | |
| Min–Max | 18–31.5 | 15–27 | 42–60 | 9–15 | 18–27 | |
| Sporangium height | n/a | n/a | n/a | n/a | ||
| Average | 44.7 ± 5.6 | 59.4 ± 4.0 | ||||
| Min–Max | 36–48 | 54–66 | ||||
| Thallus | ||||||
| Growth forms | Crustose | Decumbent | Decumbent | Prostrate, Crustose | Prostrate, Decumbent, Crustose | |
| Colors | Dark brown | Yellow–brown | Light brown | Greenish to light brown with grey band | Dark brown to orange brown | |
2.3. Interactions between Lobophora and Corals
3. Discussion
4. Materials and Methods
4.1. Sampling Sites
4.2. DNA Extraction and Phylogenetic Analyses
4.3. Morphological and Anatomical Characteristics
4.4. Ecological Studies
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vieira, C. Lobophora-coral interactions and phase shifts: Summary of current knowledge and future directions. Aquat. Ecol. 2020, 54, 1–20. [Google Scholar] [CrossRef]
- Vieira, C.; D’hondt, S.; De Clerck, O.; Payri, C.E. Toward an inordinate fondness for stars, beetles and Lobophora? Species diversity of the genus Lobophora (Dictyotales, Phaeophyceae) in New Caledonia. J. Phycol. 2014, 50, 1101–1119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, N.E.; Lane, C.E.; Le Gall, L.; Gey, D.; Bigney, A.R.; De Reviers, B.; Rousseau, F.; Schneider, C.W. A barcode analysis of the genus Lobophora (Dictyotales, Phaeophyceae) in the western Atlantic Ocean with four novel species and the epitypification of L. variegata (J.V. Lamouroux) E.C. Oliveira. Eur. J. Phycol. 2015, 50, 481–500. [Google Scholar] [CrossRef] [Green Version]
- Vieira, C.; Camacho, O.; Wynne, M.J.; Mattio, L.; Anderson, R.; Bolton, J.J.; Sansón, M.; D’Hondt, S.; Leliaert, F.; Fredericq, S.; et al. Shedding new light on old algae: Matching names and sequences in the brown algal genus Lobophora (Dictyotales, Phaeophyceae). Taxon 2016, 65, 689–707. [Google Scholar] [CrossRef] [Green Version]
- Vieira, C.; Camacho, O.; Sun, Z.; Fredericq, S.; Leliaert, F.; Payri, C.; De Clerck, O. Historical biogeography of the highly diverse brown seaweed Lobophora (Dictyotales, Phaeophyceae). Mol. Phylogenet. Evol. 2017, 110, 81–92. [Google Scholar] [CrossRef] [Green Version]
- Camacho, O.; Fernández-García, C.; Vieira, C.; Gurgel, C.F.D.; Norris, J.N.; Freshwater, D.W.; Fredericq, S. The systematics of Lobophora (Dictyotales, Phaeophyceae) in the Western Atlantic and Eastern Pacific Oceans: Eight new species. J. Phycol. 2019, 55, 611–624. [Google Scholar] [CrossRef]
- Vieira, C.; De Clerck, O.; Millet, L.; Payri, C.E. Description of ten new Lobophora species from the Bismarck Sea (Papua New Guinea). Phycol. Res. 2019, 67, 228–238. [Google Scholar] [CrossRef]
- Vieira, C.; Morrow, K.; D’Hondt, S.; Camacho, O.; Engelen, A.H.; Payri, C.E.; De Clerck, O. Divesity, ecology, biogeography, and evolution of the prevalent brown algal genus Lobophora in the Greater Caribbean Sea, including the description of five new species. J. Phycol. 2020, 56, 592–607. [Google Scholar] [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. 2022. Available online: http://www.algaebase.org (accessed on 22 August 2022).
- Vieira, C.; Payri, C.; De Clerck, O. Overgrowth and killing of corals by the brown alga Lobophora hederacea (Dictyotales, Phaeophyceae) on healthy reefs in New Caledonia: A new case of the epizoism syndrome. Phycol. Res. 2015, 63, 152–153. [Google Scholar] [CrossRef]
- Jompa, J.; McCook, L.J. Effects of competition and herbivory on interactions between a hard coral and a brown alga. J. Exp. Mar. Biol. Ecol. 2002, 271, 25–39. [Google Scholar] [CrossRef]
- Mumby, P.J.; Foster, N.L.; Fahy, E.A.G. Patch dynamics of coral reef macroalgae under chronic and acute disturbance. Coral Reefs 2005, 24, 681–692. [Google Scholar] [CrossRef]
- Nugues, M.; Bak, R. Long-term dynamics of the brown macroalga Lobophora variegata on deep reefs in Curaçao. Coral Reefs 2008, 27, 389–393. [Google Scholar] [CrossRef]
- Lesser, M.P.; Slattery, M. Phase shift to algal dominated communities at mesophotic depths associated with lionfish (Pterois volitans) invasion on a Bahamian coral reef. Biol. Invasions 2011, 13, 1855–1868. [Google Scholar] [CrossRef]
- Longo, G.; Hay, M. Does seaweed–coral competition make seaweeds more palatable? Coral Reefs 2014, 34, 87–96. [Google Scholar] [CrossRef]
- Titlyanov, E.A.; Titlyanova, T.V.; Arvedlund, M. Finding the winners in competition for substratum between coral polyps and epilithic algae on damaged colonies of the coral Porites lutea. Mar. Biodivers. Rec. 2009, 2, e85. [Google Scholar] [CrossRef]
- Roff, G.; Doropoulos, C.; Zupan, M.; Rogers, A.; Steneck, R.S.; Golbuu, Y.; Mumby, P.J. Phase shift facilitation following cyclone disturbance on coral reefs. Oecologia 2015, 178, 1193–1203. [Google Scholar] [CrossRef] [PubMed]
- Fong, J.; Deignan, L.K.; Bauman, A.G.; Steinberg, P.D.; McDougald, D.; Todd, P.A. Contact- and Water-Mediated Effects of Macroalgae on the Physiology and Microbiome of Three Indo-Pacific Coral Species. Front. Mar. Sci. 2020, 6, 831. [Google Scholar] [CrossRef]
- Briones-Fourzán, P.; Lozano-Álvarez, E. The importance of Lobophora variegata (Phaeophyta: Dictyotales) as a habitat for small juveniles of Panulirus argus in a tropical reef lagoon. Bull. Mar. Sci. 2001, 68, 207–219. [Google Scholar]
- Sangil, C.; Sanson, M.; Afonso-Carrillo, J. Spatial variation patterns of subtidal seaweed assemblages along a subtropical oceanic archipelago: Thermal gradient vs herbivore pressure. Estuar. Coast. Shelf Sci. 2011, 94, 322–333. [Google Scholar] [CrossRef]
- Granados-Cifuentes, C.; Neigel, J.; Leberg, P.; Rodriguez-Lanetty, M. Genetic diversity of free-living Symbiodinium in the Caribbean: The importance of habitats and seasons. Coral Reefs 2015, 34, 927–939. [Google Scholar] [CrossRef]
- Vieira, C.; Stenger, P.L.; Moleana, T.; De Clerck, O.; Payri, C. Limited interspecific variation in grazing susceptibility of the brown alga Lobophora to herbivory. J. Exp. Mar. Biol. Ecol. 2019, 518, 151175. [Google Scholar] [CrossRef]
- Rasher, D.B.; Hay, M.E. Seaweed allelopathy degrades the resilience and function of coral reefs. Commun. Integr. Biol. 2010, 3, 564–566. [Google Scholar] [CrossRef]
- Slattery, M.; Lesser, M.P. Allelopathy in the tropical alga Lobophora variegata (Phaeophyceae): Mechanistic basis for a phase shift on mesophotic coral reefs? J. Phycol. 2014, 50, 493–505. [Google Scholar] [CrossRef]
- Vieira, C.; Thomas, O.P.; Culioli, G.; Genta-Jouve, G.; Houlbreque, F.; Gaubert, J.; De Clerck, O.; Payri, C.E. Allelopathic interactions between the brown algal genus Lobophora (Dictyotales, Phaeophyceae) and scleractinian corals. Sci. Rep. 2016, 6, 18637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morrow, K.M.; Bromhall, K.; Motti, C.A.; Munn, C.B.; Bourne, D.G. Allelochemicals produced by brown macroalgae of the Lobophora genus are active against coral larvae and associated bacteria, supporting pathogenic shifts to vibrio dominance. Appl. Environ. Microbiol. 2017, 83, e02391-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fong, J.; Lim, Z.W.; Bauman, A.G.; Valiyaveettil, S.; Liao, L.M.; Yip, Z.T.; Todd, P.A. Allelopathic effects of macroalgae on Pocillopora acuta coral larvae. Mar. Environ. Res. 2019, 151, 104745. [Google Scholar] [CrossRef]
- Evensen, N.R.; Doropoulos, C.; Wong, K.J.; Mumby, P.J. Stage-specific effects of Lobophora on the recruitment success of a reef-building coral. Coral Reefs 2019, 38, 489–498. [Google Scholar] [CrossRef]
- Mayakun, J.; Prathep, A. Seasonal variations in diversity and abundance of macroalgae at Samui Island, Surat Thani Province, Thailand. Songklanakarin J. Sci. Technol. 2005, 27, 653–663. [Google Scholar]
- Coppejans, E.; Prathep, A.; Leliaert, F.; Lewmanomont, K.; De Clerck, O. Seaweeds of Mu Ko Tha Lae Tai (SE Thailand): Methodologies and Field Guide to the Dominant Species; Biodiversity Research and Training Program (BRT): Bangkok, Thailand, 2010. [Google Scholar]
- Prathep, A.; Pongparadon, S.; Darakrai, A.; Wichachucherd, B.; Sinutok, S. Diversity and distribution of seaweed at Khanom-Mu Ko Thale Tai National Park, Nakhon Si Thammarat Province, Thailand. Songklanakarin J. Sci. Technol. 2011, 33, 633–640. [Google Scholar]
- Kaewsuralikhit, C.; Duangdee, T. Marine Algae at Hin-Grood Reef and Koh Talu Islands, Prachuab Khiri Khan Province. KKU Sci. J. 2012, 40, 135–142. [Google Scholar]
- Coppejans, E.; Prathep, A.; Lewmanomont, K.; Hayashizaki, K.; De Clerck, O.; Leliaert, F.; Terada, R. Seaweeds and Seagrasses of the Southern Andaman Sea Coast of Thailand; The Kagoshima University Museum: Kagoshima, Japan, 2017. [Google Scholar]
- Vieira, C.; Rasoamanendrika, F.A.; Zubia, M.; Bolton, J.J.; Anderson, R.J.; Engelen, A.H.; D’hondt, S.; Leliaert, F.; Payri, C.; Kawai, H.; et al. Lobophora (Dictyotales, Phaeophyceae) from the western Indian Ocean: Diversity and biogeography. S. Afr. J. Bot. 2021, 142, 230–246. [Google Scholar] [CrossRef]
- Alejandre, J.G. Interactions between Macroalgae and Corals on Coral Reefs around Koh Tao, Gulf of Thailand; University of Amsterdam: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Titioatchasai, J.; Prathep, A.; Mayakun, J. Pattern of algal succession in the tropical subtidal coral reef community at Koh Taen, Mu Ko Thale Tai National Park, the Gulf of Thailand. J. Fish. Environ. 2019, 43, 11–18. [Google Scholar]
- Müller, M.; Staab, C.F.K.; Puk, L.D.; Schoenig, E.M.; Ferse, S.C.A.; Wild, C. The rabbitfish Siganus virgatus as key macroalgae browser in coral reefs of the Gulf of Thailand. Diversity 2021, 13, 123. [Google Scholar] [CrossRef]
- Kwan, V.; Yip, Z.T.; Fong, J.; Huang, D. Diversity and phylogeny of the brown alga Lobophora (Dictyotales, Phaeophyceae) in Singapore. Phytotaxa 2021, 496, 215–227. [Google Scholar] [CrossRef]
- Diaz-Pulido, G.; McCook, L.J. Effects of live coral, epilithic algal communities and substrate type on algal recruitment. Coral Reefs 2004, 23, 225–233. [Google Scholar] [CrossRef]
- Puk, L.D.; Vieira, C.; Roff, G.; De Clerck, O.; Mumby, P.J. Cryptic diversity in the macroalgal genus Lobophora (Dictyotales) reveals environmental drivers of algal assemblages. Mar. Biol. 2020, 167, 188. [Google Scholar] [CrossRef]
- Bennett, S.; Verge´s, A.; Bellwood, D. Branching coral as a macroalgal refuge in a marginal coral reef system. Coral Reefs 2010, 29, 471–480. [Google Scholar] [CrossRef]
- Aunkhongthong, W.; Yeemin, T.; Rangseethampanya, P.; Chamchoy, C.; Ruengthong, C.; Samsuvan, W.; Thummasan, M.; Sutthacheep, M. Coral community structures on shallow reef flat, reef slope and underwater pinnacles in Mu Ko Chumphon, the western Gulf of Thailand. RIST 2021, 4, 1–7. [Google Scholar]
- Sutthacheep, M.; Chamchoy, C.; Pengsakun, S.; Klinthong, W.; Yeemin, T. Assessing the resilience potential of inshore and offshore coral communities in the western Gulf of Thailand. J. Mar. Sci. Eng. 2019, 7, 408. [Google Scholar] [CrossRef] [Green Version]
- Cheal, A.J.; Emslie, M.J.; Macneil, M.A.; Miller, I.; Sweatman, H. Spatial variation in the functional characteristics of herbivorous fish communities and the resilience of coral reefs. Ecol. Appl. 2013, 23, 174–188. [Google Scholar] [CrossRef] [Green Version]
- Wenger, A.; Fabricius, K.E.; Jones, G.P.; Brodie, J.E. Sedimentation, eutrophication and chemical pollution: Effects on coral reef fishes. In Coral Reef Fishes; Mora, C., Ed.; Cambridge University Press: Cambridge, UK, 2015. [Google Scholar]
- Puk, L.D.; Marshell, A.; Dwyer, J.; Evensen, N.R.; Mumby, P.J. Refuge-dependent herbivory controls a key macroalga on coral reefs. Coral Reefs 2020, 39, 953–965. [Google Scholar] [CrossRef]
- Coen, L.D.; Tanner, C.E. Morphological variation and differential susceptibility to herbivory in the tropical brown alga Lobophora variegata. Mar. Ecol. Prog. Ser. 1989, 54, 287–298. [Google Scholar] [CrossRef]
- Verge´s, A.; Vanderklift, M.A.; Doropoulos, C.; Hyndes, G.A. Spatial patterns in herbivory on a coral reef are influenced by structural complexity but not by algal traits. PLoS ONE 2011, 6, e17115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Hanyuda, T.; Lim, P.E.; Tanaka, J.; Gurgel, C.F.D.; Kawai, H. Taxonomic revision of the genus Lobophora (Dictyotales, Phaeophyceae) based on morphological evidence and analyses rbcL and cox3 gene sequences. Phycologia 2012, 51, 500–512. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate Maximum-Likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [PubMed]
- English, S.; Wilkinson, C.; Baker, V. Survey Manual for Tropical Marine Resouces; Australian Institute of Marine Science: Queensland, Australia, 1997. [Google Scholar]
- Veron, J.E.N. Corals of the World; Australian Institute of Marine Science: Townsville, Australia, 2000. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 29 August 2022).
- Bittner, L.; Payri, C.; Couloux, A.; Cruaud, C.; De Reviers, B.; Rousseau, F. Molecular phylogeny of the Dictyotales and their position within the Phaeophyceae, based on nuclear, plastid and mitochondrial DNA sequence data. Mol. Phylogenet. Evol. 2008, 49, 211–226. [Google Scholar]
- Draisma, S.G.; Prud’homme van Reine, W.F.; Stam, W.T.; Olsen, J.L. A reassessment of phylogenetic relationships within the Phaeophyceae based on RUBISCO large subunit and ribosomal DNA sequences. J. Phycol. 2001, 37, 586–603. [Google Scholar]
- Win, N.-N.; Hanyuda, T.; Arai, S.; Uchimura, M.; Aboott, I.A.; Kawai, H. Three new records of Padina in Japan based on morphological and molecular markers. Phycol. Res. 2008, 56, 288–300. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klomjit, A.; Vieira, C.; Mattos, F.M.G.; Sutthacheep, M.; Sutti, S.; Kim, M.-S.; Yeemin, T. Diversity and Ecology of Lobophora Species Associated with Coral Reef Systems in the Western Gulf of Thailand, including the Description of Two New Species. Plants 2022, 11, 3349. https://doi.org/10.3390/plants11233349
Klomjit A, Vieira C, Mattos FMG, Sutthacheep M, Sutti S, Kim M-S, Yeemin T. Diversity and Ecology of Lobophora Species Associated with Coral Reef Systems in the Western Gulf of Thailand, including the Description of Two New Species. Plants. 2022; 11(23):3349. https://doi.org/10.3390/plants11233349
Chicago/Turabian StyleKlomjit, Anirut, Christophe Vieira, Felipe M. G. Mattos, Makamas Sutthacheep, Suttikarn Sutti, Myung-Sook Kim, and Thamasak Yeemin. 2022. "Diversity and Ecology of Lobophora Species Associated with Coral Reef Systems in the Western Gulf of Thailand, including the Description of Two New Species" Plants 11, no. 23: 3349. https://doi.org/10.3390/plants11233349