
An Overview of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae), an Emerging Cause of Human Angiostrongylosis on the Indian Subcontinent



Abstract
:1. Introduction
2. Biology and Life Cycle of Angiostrongylus cantonensis
3. Epidemiology and Clinical Manifestations of Human Angiostrongylosis in the Indian Subcontinent

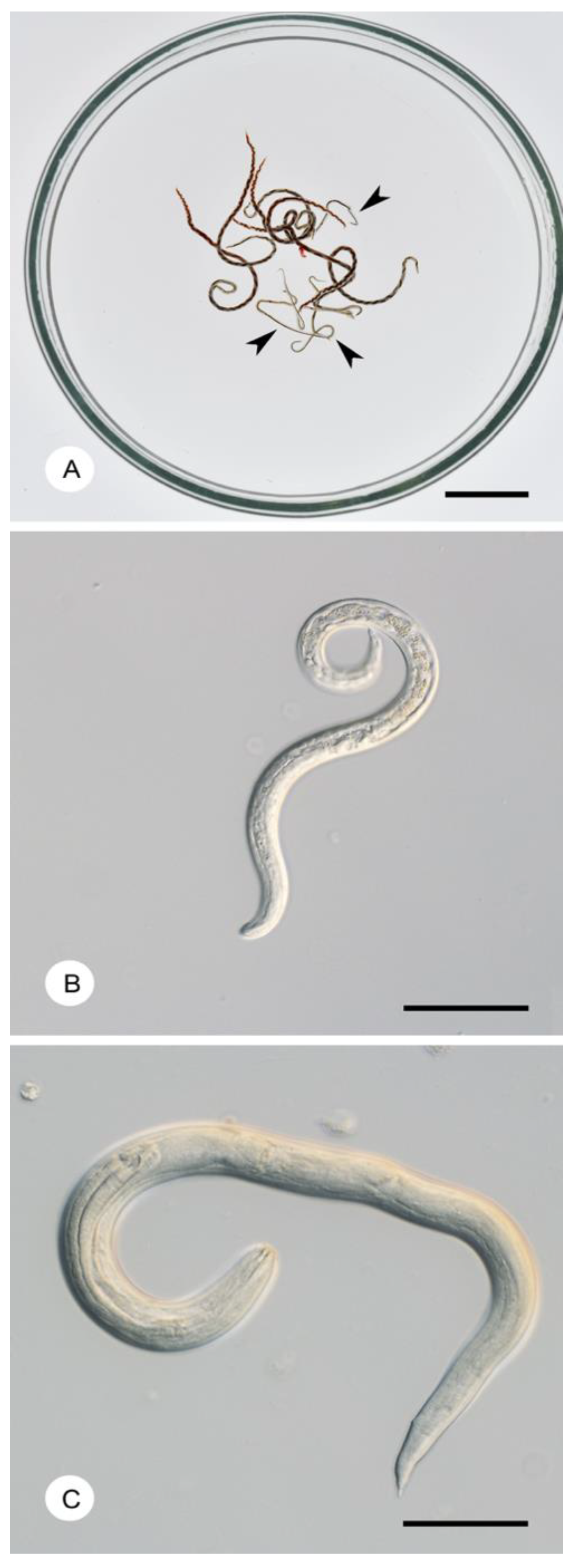{kind=link}
{kind=link}
{kind=link}
| Country: State | EM Cases | OC Cases | Diagnostics Method | Monitor Lizard Consumption | References |
|---|---|---|---|---|---|
| India: Delhi | 1 | Slit lamp examination | [43] | ||
| India: Delhi | 1 | Clinical examination, ultrasound B scan, and fundus fluorescein angiography | [42] | ||
| India: Karnataka | 1 | History of raw monitor lizard meat consumption, clinical presentation, and EE in CSF | + | [64] | |
| India: Karnataka | 1 | Serum and CSF antibodies to A. cantonensis 31-kDa antigen positive and magnetic resonance imaging (MRI) | + | [63] | |
| India: Karnataka | 1 | MRI findings, CSF examination, larvae in CSF wet mount | + | [44] | |
| India: Kerala | 5 | History of raw monitor lizard meat consumption, clinical presentation, and EE in CSF | + | [66] | |
| India: Kerala | 10 | History of raw monitor lizard meat consumption, clinical presentation, EE in CSF., larvae in CSF wet mount, MRI | + | [48] | |
| India: Kerala | 3 | EE in CSF and peripheral blood | [62] | ||
| India: Kerala | 1 * | Ophthalmological examination, EE in CSF, and microscopic examination of the worm retrieved from eye | [36] | ||
| India: Kerala | 1 | EE in CSF, real-time PCR for A. cantonensis | [46] | ||
| India: Maharashtra | 1 | Not mentioned | [6] | ||
| India: Maharashtra | 2 | History of raw slug consumption and EE in CSF | [68] | ||
| India: Maharashtra | 2 | EE in CSF and, peripheral blood | ** | [47] | |
| India: Maharashtra | 1 | Histological examination of brain tissue after autopsy | [65] | ||
| India: Madhya Pradesh | 1 | Slit lamp examination and anterior segment optical coherence topography (AS-OCT) | [38] | ||
| India: Madhya Pradesh | 1 | Slit lamp examination and microscopic examination of the worm retrieved from eye | [39] | ||
| India: Puducherry | 1 | History of raw monitor lizard meat consumption, clinical presentation, EE in CSF analysis | + | [67] | |
| India: Telangana | 1 | Computerized tomography (CT) scans, parietal craniotomy examination of the worm retrieved from cerebral abscess | [69] | ||
| India: West Bengal | 1 | Microscopic examination of the worm retrieved from eye | [6] | ||
| Sri Lanka | 1 | Ophthalmological examination and microscopic examination of the worm retrieved from eye | [70,71] | ||
| Sri Lanka | 1 | Ophthalmological examination and microscopic examination of the worm retrieved from eye | [72] | ||
| Sri Lanka | 1 | Fundoscopic ophthalmological examination and microscopic examination of the worm retrieved from eye | [73] | ||
| Sri Lanka | 1 | EE in CSF and peripheral blood | [74] | ||
| Sri Lanka | 1 | Ophthalmological examination | [75] | ||
| Sri Lanka | 1 | History of raw monitor lizard meat consumption, clinical presentation, EE in CSF | + | [45] | |
| Sri Lanka | 1 | Fundoscopic ophthalmological examination and microscopic examination of the worm retrieved from eye | [76] | ||
| Nepal: Kathmandu | 1 | Slit lamp examination, microscopic examination of the worm retrieved from eye | [77] | ||
| Nepal | 1 | Ophthalmological examination and microscopic examination of the worm retrieved from eye | [78] |
4. Global Distribution and Prevalence of Angiostrongylus cantonensis in the Indian Subcontinent
5. Angiostrongylus cantonensis in the Definitive Rodent Host
5.1. Angiostrongylus cantonensis in Intermediate Hosts
5.2. Snail Consumption
6. Angiostrongylus cantonensis in Paratenic Reptilian and Amphibian Hosts in the Context of Local Consumption in India
6.1. Monitor Lizards (Varanidae)
6.2. Snakes
6.3. Amphibians
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Morgan, E.R.; Modry, D.; Paredes-Esquivel, C.; Foronda, P.; Traversa, D. Angiostrongylosis in animals and humans in Europe. Pathogens 2021, 10, 1236. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-P.; Lai, D.-H.; Zhu, X.-Q.; Chen, X.-G.; Lun, Z.-R. Human angiostrongyliasis. Lancet Infect. Dis. 2008, 8, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Eamsobhana, P. Angiostrongyliasis in Thailand: Epidemiology and laboratory investigations. Hawaii J. Med. Public Health 2013, 72, 28–32. [Google Scholar] [PubMed]
- Punyagupta, S.; Bunnag, T.; Juttijudata, P.; Rosen, L. Eosinophilic meningitis in Thailand. Epidemiologic studies of 484 typical cases and the etiologic role of Angiostrongylus cantonensis. Am. J. Trop. Med. Hyg. 1970, 19, 950–958. [Google Scholar] [CrossRef] [PubMed]
- Watthanakulpanich, D.; Jakkul, W.; Chanapromma, C.; Ketboonlue, T.; Dekumyoy, P.; Lv, Z.; Chan, A.H.E.; Thaenkham, U.; Chaisiri, K. Co-occurrence of Angiostrongylus malaysiensis and Angiostrongylus cantonensis DNA in cerebrospinal fluid: Evidence from human eosinophilic meningitis after ingestion of raw snail dish in Thailand. Food Waterborne Parasitol. 2021, 24, e00128. [Google Scholar] [CrossRef]
- Parmeter, S.N.; Chowdhury, A.B. Angiostrongylus cantonensis in India. Bull. Calcutta Sch. Trop. Med. 1966, 14, 38. [Google Scholar]
- Turck, H.C.; Fox, M.T.; Cowie, R.H. Paratenic hosts of Angiostrongylus cantonensis and their relation to human neuroangiostrongyliasis globally. One Health 2022, 15, 100426. [Google Scholar] [CrossRef]
- Lecompte, E.; Aplin, K.; Denys, C.; Catzeflis, F.; Chades, M.; Chevret, P. Phylogeny and biogeography of African Murinae based on mitochondrial and nuclear gene sequences, with a new tribal classification of the subfamily. BMC Evol. Biol. 2008, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.-T. Un Nouveau nématode pulmonaire, Pulmonema cantonensis, n.g., n. sp. Ann. Parasitol. Hum. Comp. 1935, 13, 312–317. [Google Scholar] [CrossRef] [Green Version]
- Alicata, J.E. Biology and distribution of the Rat lungworm, Angiostrongylus cantonensis, and its relationship to eosinophilic meningoencephalitis and other neurological disorders of man and animals. Adv. Parasitol. 1965, 3, 223–248. [Google Scholar] [CrossRef]
- Walden, H.D.S.; Slapcinsky, J.D.; Roff, S.; Calle, J.M.; Goodwin, Z.D.; Stern, J.; Corlett, R.; Conway, J.; McIntosh, A. Geographic distribution of Angiostrongylus cantonensis in wild rats (Rattus rattus) and terrestrial snails in Florida, USA. PLoS ONE 2017, 12, e0177910. [Google Scholar] [CrossRef] [Green Version]
- Wallace, G.D.; Rosen, L. Studies on eosinophilic meningitis: I. Observations on the geographic distribution of Angiostrongylus cantonensis in the Pacific area and its prevalence in wild rats. Am. J. Epidemiol. 1965, 81, 52–62. [Google Scholar] [CrossRef]
- Mackerras, M.; Sandars, D. The life history of the rat lung-worm, Angiostrongylus cantonensis (Chen) (Nematoda: Metastrongylidae). Aust. J. Zool. 1955, 3, 1–21. [Google Scholar] [CrossRef]
- Červená, B.; Modrý, D.; Fecková, B.; Hrazdilová, K.; Foronda, P.; Alonso, A.M.; Lee, R.; Walker, J.; Niebuhr, C.N.; Malik, R.; et al. Low diversity of Angiostrongylus cantonensis complete mitochondrial DNA sequences from Australia, Hawaii, French Polynesia and the Canary Islands revealed using whole genome next-generation sequencing. Parasites Vectors 2019, 12, 241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alicata, J. The discovery of Angiostrongylus cantonensis as a cause of human eosinophilic meningitis. Parasitol. Today 1991, 7, 151–153. [Google Scholar] [CrossRef]
- Mason, K.V.; Prescott, C.W.; Kelly, W.R.; Waddell, A.H. Granulomatous encephalomyelitis of puppies due to Angiostrongylus cantonensis. Aust. Vet. J. 1976, 52, 295. [Google Scholar] [CrossRef]
- Wright, J.; Kelly, W.; Hamilton, J.; Wadell, A. Equine neural angiostrongylosis. Aust. Veter J. 1991, 68, 58–60. [Google Scholar] [CrossRef]
- Carlisle, M.S.; Prociv, P.; Grennan, J.; Pass, M.A.; Campbell, G.L.; Mudie, A. Cerebrospinal angiostrongyliasis in five captive tamarins (Sanguinus spp.). Aust. Veter J. 1998, 76, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.; Carlisle, M.; Prociv, P. Neuro-angiostrongylosis in wild Black and Grey-headed flying foxes (Pteropus spp.). Aust. Vet. J. 2002, 80, 554–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mason, K.V. Canine neural angiostrongylosis: The clinical and therapeutic features of 55 natural cases. Aust. Vet. J. 1987, 64, 201–203. [Google Scholar] [CrossRef]
- Lunn, J.A.; Lee, R.; Smaller, J.; MacKay, B.M.; King, T.; Hunt, G.B.; Martin, P.; Krockenberger, M.B.; Spielman, D.; Malik, R. Twenty two cases of canine neural angiostrongylosis in eastern Australia (2002–2005) and a review of the literature. Parasites Vectors 2012, 5, 70. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.R.; Hayes, K.A.; Yeung, N.W.; Cowie, R.H. Diverse gastropod hosts of Angiostrongylus cantonensis, the rat lungworm, globally and with a focus on the Hawaiian Islands. PLoS ONE 2014, 9, e94969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiengo, S.C.; Simões, R.D.O.; Fernandez, M.A.; Maldonado, A. Angiostrongylus cantonensis and rat lungworm disease in Brazil. Hawaii J. Med. Public Health 2013, 72, 18–22. [Google Scholar]
- Howe, K.; Kaluna, L.; Lozano, A.; Fischer, B.T.; Tagami, Y.; McHugh, R.; Jarvi, S. Water transmission potential of Angiostrongylus cantonensis: Larval viability and effectiveness of rainwater catchment sediment filters. PLoS ONE 2019, 14, e0209813. [Google Scholar] [CrossRef] [Green Version]
- Modrý, D.; Fecková, B.; Putnová, B.; Manalo, S.M.; Otranto, D. Alternative pathways in Angiostrongylus cantonensis (Metastrongyloidea: Angiostrongylidae) transmission. Parasitology 2021, 148, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Diao, Z.; Wang, J.; Qi, H.; Li, X.; Zheng, X.; Yin, C. Human ocular angiostrongyliasis: A literature review. Trop. Doct. 2011, 41, 76–78. [Google Scholar] [CrossRef] [PubMed]
- Beaver, P.C.; Rosen, L. Memorandum on the first report of Angiostrongylus in man, by Nomura and Lin, 1945. Am. J. Trop. Med. Hyg. 1964, 13, 589–590. [Google Scholar] [CrossRef]
- Ansdell, V.; Wattanagoon, Y. Angiostrongylus cantonensis in travelers: Clinical manifestations, diagnosis, and treatment. Curr. Opin. Infect. Dis. 2018, 31, 399–408. [Google Scholar] [CrossRef]
- Kwon, E.; Ferguson, T.M.; Park, S.Y.; Manuzak, A.; Qvarnstrom, Y.; Morgan, S.; Ciminera, P.; Murphy, G.S. A severe case of Angiostrongylus eosinophilic meningitis with encephalitis and neurologic sequelae in Hawa’i. Hawaii J. Med. Public Health 2013, 72, 41–45. [Google Scholar]
- Murphy, G.S.; Johnson, S. Clinical aspects of eosinophilic meningitis and meningoencephalitis caused by Angiostrongylus cantonensis, the rat lungworm. Hawaii J. Med. Public Health 2013, 72, 35. [Google Scholar]
- Feng, Y.; Nawa, Y.; Sawanyavisuth, K.; Lv, Z.; Wu, Z.-D. Comprehensive review of ocular angiostrongyliasis with special reference to optic neuritis. Korean J. Parasitol. 2013, 51, 613–619. [Google Scholar] [CrossRef]
- Kumar, V.; Kyprianou, I.; Keenan, J.M. Ocular angiostrongyliasis: Removal of a live nematode from the anterior chamber. Eye 2004, 19, 229–230. [Google Scholar] [CrossRef] [Green Version]
- Mattis, A.; Mowatt, L.; Lue, A.; Lindo, J.; Vaughan, H. Ocular angiostrongyliasis—First case report from Jamaica. West Indian Med. J. 2009, 58, 383–385. [Google Scholar] [PubMed]
- McCarthy, K.; Liu, K.; Kokame, G.T.; Merrill, P.T.; Gilca, M.; Cohen, J. First case of subretinal ocular angiostrongyliasis associated with retinal detachment in the United States. Hawaii J. Health Soc. Welf. 2021, 80, 40. [Google Scholar] [PubMed]
- Sinawat, S.; Trisakul, T.; Choi, S.; Morley, M.; Sinawat, S.; Yospaiboon, Y. Ocular angiostrongyliasis in Thailand: A retrospective analysis over two decades. Clin. Ophthalmol. 2019, 13, 1027–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baheti, N.N.; Sreedharan, M.; Krishnamoorthy, T.; Nair, M.D.; Radhakrishnan, K. Eosinophilic meningitis and an ocular worm in a patient from Kerala, South India. Case Rep. 2009, 2009, bcr2007122093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawanyawisuth, K.; Kitthaweesin, K.; Limpawattana, P.; Intapan, P.; Tiamkao, S.; Jitpimolmard, S.; Chotmongkol, V. Intraocular angiostrongyliasis: Clinical findings, treatments and outcomes. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 497–501. [Google Scholar] [CrossRef]
- Sen, P.; Date, A.; Jain, E.; Mohan, A. Angiostrongylus—A technique for removing a rare parasite from the cornea of a child. J. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2019, 23, 119–120. [Google Scholar] [CrossRef]
- Aishwarya, A.; Tiwari, U.S.; Gandhi, S.; Sisodia, P. Angiostrongylus cantonensis in anterior chamber. Indian J. Ophthalmol. 2019, 67, 158–160. [Google Scholar] [CrossRef]
- Renapurkar, D.M.; Bhopale, M.K.; Limaye, L.S.; Sharma, K.D. Prevalence of Angiostrongylus cantonensis infection in commensal rats in Bombay. J. Helminthol. 1982, 56, 345–349. [Google Scholar] [CrossRef]
- Thomas, M.; Thangavel, M.; Thomas, R.P. Angiostrongylus cantonensis (Nematoda, Metastrongylidae) in bandicoot rats in Kerala, South India. Infect. Dis. 2015, 8, 324–326. [Google Scholar]
- Mehta, D.K.; Arora, R.; Chauhan, D.; Shroff, D.; Narula, R. Chemo-paralysis for the removal of a live intraocular worm in ocular angiostrongyliasis. Clin. Exp. Ophthalmol. 2006, 34, 493–495. [Google Scholar] [CrossRef]
- Malhotra, S.; Mehta, D.K.; Arora, R.; Chauhan, D.; Ray, S.; Jain, M. Ocular angiostrongyliasis in a child—First case report from India. J. Trop. Pediatr. 2006, 52, 223–225. [Google Scholar] [CrossRef] [Green Version]
- Rai, S.; Madi, D.; Pai, S.; Baliga, S. Unusual larva in the CSF and unique MRI findings in a case of eosinophilic meningitis. J. Clin. Imaging Sci. 2014, 4, 76. [Google Scholar] [CrossRef]
- Hidelaratchi, M.D.P.; Riffsy, M.T.M.; Wijesekera, J.C. A case of eosinophilic meningitis following monitor lizard meat consumption, exacerbated by anthelminthics. Ceylon Med. J. 2010, 50, 84–86. [Google Scholar] [CrossRef] [Green Version]
- Raveendrakumar, A.G.; Remadevi, G.S.; Eapen, E.K.; Vijayamma, A.K.T. Eosinophilic meningitis in a toddler. Indian Pediatr. 2021, 58, 187–188. [Google Scholar] [CrossRef]
- Shah, I.; Barot, S.; Madvariya, M. Eosinophilic meningitis: A case series and review of literature of Angiostrongylus cantonensis and Gnathostoma spinigerum. Indian J. Med. Microbiol. 2015, 33, 154–158. [Google Scholar] [CrossRef] [PubMed]
- Parameswaran, K. Case series of eosinophilic meningoencephalitis from South India. Ann. Indian Acad. Neurol. 2006, 9, 217. [Google Scholar] [CrossRef]
- Ianas, V.; Kuberski, T. Eosinophilic meningitis due to Angiostrongylus cantonensis. Infect. Med. 2008, 25, 366–368. [Google Scholar]
- Wang, Q.-P.; Wu, Z.-D.; Wei, J.; Owen, R.L.; Lun, Z.-R. Human Angiostrongylus cantonensis: An update. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 389–395. [Google Scholar] [CrossRef]
- Wilkins, P.P.; Qvarnstrom, Y.; Whelen, A.C.; Saucier, C.; Da Silva, A.J.; Eamsobhana, P. The current status of laboratory diagnosis of Angiostrongylus cantonensis infections in humans using serologic and molecular methods. Hawaii J. Med. Public Health 2013, 72, 55–57. [Google Scholar]
- de Melo, L.C.V.; de Souza, F.C.R.; Baccin, A.D.O.; Mota, D.J.G.D.; Pereira-Chioccola, V.L.; Pinto, P.L.S. Immunoanalysis of different antigenic preparations of Angiostrongylus cantonensis for neuroangiostrongyliasis diagnosis improvement. Mem. Inst. Oswaldo Cruz 2022, 117, e220086. [Google Scholar] [CrossRef]
- Qvarnstrom, Y.; Sullivan, J.J.; Bishop, H.S.; Hollingsworth, R.; Da Silva, A.J. PCR-based detection of Angiostrongylus cantonensis in tissue and mucus secretions from molluscan hosts. Appl. Environ. Microbiol. 2007, 73, 1415–1419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qvarnstrom, Y.; Xayavong, M.; Chea, N.; da Silva, A.C.A.; Heng, S.; Park, S.Y.; Calimlim, P.S.; Johnson, S.; Whelen, A.C.; Fox, L.M.; et al. Real-time polymerase chain reaction detection of Angiostrongylus cantonensis DNA in cerebrospinal fluid from patients with eosinophilic meningitis. Am. J. Trop. Med. Hyg. 2016, 94, 176–181. [Google Scholar] [CrossRef] [Green Version]
- Maleewong, W.; Sombatsawat, P.; Intapan, P.M.; Wongkham, C.; Chotmongkol, V. Immunoblot evaluation of the specificity of the 29-kDa antigen from young adult female worms Angiostrongylus cantonensis for immunodiagnosis of human angiostrongyliasis. Asian Pac. J. Allergy Immunol. 2001, 19, 267. [Google Scholar]
- Eamsobhana, P.; Gan, X.; Ma, A.; Wang, Y.; Wanachiwanawin, D.; Yong, H. Dot immunogold filtration assay (DIGFA) for the rapid detection of specific antibodies against the rat lungworm Angiostrongylus cantonensis (Nematoda: Metastrongyloidea) using purified 31-kDa antigen. J. Helminthol. 2014, 88, 396–401. [Google Scholar] [CrossRef]
- Chen, J.-X.; Chen, M.-X.; Ai, L.; Chen, J.-H.; Chen, S.-H.; Zhang, Y.-N.; Cai, Y.-C.; Zhu, X.-Q.; Zhou, X.-N. A protein microarray for the rapid screening of patients suspected of infection with various food-borne helminthiases. PLoS Negl. Trop. Dis. 2012, 6, e1899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barratt, J.; Chan, D.; Sandaradura, I.; Malik, R.; Spielman, D.; Lee, R.; Marriott, D.; Harkness, J.; Ellis, J.; Stark, D. Angiostrongylus cantonensis: A review of its distribution, molecular biology and clinical significance as a human pathogen. Parasitology 2016, 143, 1087–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.-Y.; Song, H.-Q.; Zhang, R.-L.; Chen, M.-X.; Xu, M.-J.; Ai, L.; Chen, X.-G.; Zhan, X.-M.; Liang, S.-H.; Yuan, Z.-G.; et al. Specific detection of Angiostrongylus cantonensis in the snail Achatina fulica using a loop-mediated isothermal amplification (LAMP) assay. Mol. Cell. Probes 2011, 25, 164–167. [Google Scholar] [CrossRef]
- Sears, W.J.; Qvarnstrom, Y.; Nutman, T.B. RPAcan3990: An ultrasensitive recombinase polymerase assay to detect Angiostrongylus cantonensis DNA. J. Clin. Microbiol. 2021, 59, e01185-21. [Google Scholar] [CrossRef]
- Sears, W.J.; Qvarnstrom, Y.; Dahlstrom, E.; Snook, K.; Kaluna, L.; Baláž, V.; Feckova, B.; Šlapeta, J.; Modry, D.; Jarvi, S. AcanR3990 qPCR: A novel, highly sensitive, bioinformatically-informed assay to detect Angiostrongylus cantonensis Infections. Clin. Infect. Dis. 2021, 73, e1594–e1600. [Google Scholar] [CrossRef]
- Jalal, M.J.A.; Varghese, B.; Menon, M.K.; Pratap, T.; Moheen, A.; Mathew, J.; Vijayan, T.P.; Tomy, M.P. Eosinophilic meningitis: Report of three cases from a hilly area in Kerala. Ann. Indian Acad. Neurol. 2019, 22, 108–111. [Google Scholar] [CrossRef]
- Nalini, A.; Ramakrishna, A.; Dekumoy, P.; Kumar, R.; Pakdee, W.; Saini, J.; Hegde, V. Severe form of radiculo-myelo-neuropathy with meningo-encephalitis secondary to Angiostrongylus cantonensis infection: Unusual corpus callosal lesions and serial magnetic resonance imaging findings. Neurol. India 2013, 61, 414–418. [Google Scholar] [CrossRef]
- Pai, S.; Madi, D.; Achappa, B.; Mahalingam, S.; Kendambadi, R. An interesting case of eosinophilic meningitis. J. Clin. Diagn. Res. 2013, 7, 734. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, J.P.; Shenoy, A.S.; Goel, N.A. Eosinophilic meningitis due to A. Cantonensis revealed at autopsy. Indian J. Pathol. Microbiol. 2022, 65, 420. [Google Scholar] [PubMed]
- Panackel, C.; Vishad; Cherian, G.; Vijayakumar, K.; Sharma, R. Eosinophilic meningitis due to Angiostrongylus cantonensis. Indian J. Med. Microbiol. 2006, 24, 220–221. [Google Scholar] [CrossRef] [PubMed]
- Hamide, A.; Johny, J.; Kumar, A.; Kolar, V.V. Eosinophilic meningitis caused by consumption of meat of monitor lizard (Varanus bengalensis). Neurol. India 2018, 66, 1166. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.D.; Renapurkar, D.M.; Bhopale, M.K.; Nathan, J.; Boraskar, A.; Chotani, S. Study of a focus of Angiostrongylus cantonensis in Greater Bombay. Bull Haffkine 1981, 9, 38–46. [Google Scholar]
- Purohit, A.K.; Dinakar, I.; Sundaram, C.; Ratnakar, K.S. Angiostrongylus cantonensis abscess in the brain. J. Neurol. Neurosurg. Psychiatry 1991, 54, 1015–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, H.P. Clinical notes on the effects of a nematode larva in the eye. Ceylon J. Sci. Med. Sci. 1927, 1, 141. [Google Scholar]
- Nicholls, L. Note on a nematode larva removed from the eye of a man. Ceylon J. Sci. 1925, 1, 145. [Google Scholar]
- Durette-Desset, M.-C.; Chabaud, A.G.; Cassim, M.H.S.; Ismail, M.M.; Premaratne, U.N.; Abeyewickreme, W.; Dissanaike, A.S. On an infection of a human eye with Parastrongylus (=Angiostrongylus) sp. in Sri Lanka. J. Helminthol. 1993, 67, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Wariyapola, D.; Goonesinghe, N.; Priyamanna, T.H.; Fonseka, C.; Ismail, M.; Abeyewickreme, W.; Dissanaike, A. Second case of ocular parastrongyliasis from Sri Lanka. Trans. R. Soc. Trop. Med. Hyg. 1998, 92, 64–65. [Google Scholar] [CrossRef] [PubMed]
- Alibhoy, A.T.; Senanayake, B.; Fernando, M.A.; Amarasekera, H.S.; Wijesekera, J.C. A case of eosinophilic meningitis. Ceylon Med. J. 1999, 44, 173–174. [Google Scholar]
- Dissanaike, A.S.; Ihalamulla, R.L.; Naotunne, T.S.; Senarathna, T.; Withana, D.S. Third report of ocular parastrongyliasis (angiostrongyliasis) from Sri Lanka. Parassitologia 2001, 43, 95–97. [Google Scholar]
- Ilhalamulla, R.; Fernando, S.; Weerasena, K.; Cross, J.; Dissanaike, A.; Fonseka, C. A further case of Parastrongyliasis (=Angiostrongyliasis) from the eye of a patient in Sri Lanka. Ceylon J. Med. Sci. 2008, 50, 15. [Google Scholar] [CrossRef]
- Paul, A.; Pammal, A.T. Ocular parasitosis: A rare cause of hypertensive uveitis. Indian J. Ophthalmol. 2008, 56, 501–502. [Google Scholar] [CrossRef]
- Sah, R.; Khatri, A.; Kharel, R.; Kc, H.; Rabaan, A.A.; Tiwari, R.; Dhama, K.; Malik, Y.S.; Donovan, S.; Rodriguez-Morales, A.J.; et al. Case report: Management of dead intraocular helminth parasites in asymptomatic patients. Am. J. Trop. Med. Hyg. 2020, 103, 719–722. [Google Scholar] [CrossRef]
- Peng, J.; He, Z.-P.; Zhang, S.; Lun, Z.-R.; Wu, Z.-D.; Fan, C.-K.; Brown, C.L.; Cheng, P.-C.; Peng, S.-Y.; Yang, T.-B. Phylogeography of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae) in southern China and some surrounding areas. PLoS Negl. Trop. Dis. 2017, 11, e0005776. [Google Scholar] [CrossRef] [Green Version]
- Tokiwa, T.; Hashimoto, T.; Yabe, T.; Komatsu, N.; Akao, N.; Ohta, N. First report of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae) infections in invasive rodents from five islands of the Ogasawara Archipelago, Japan. PLoS ONE 2013, 8, e70729. [Google Scholar] [CrossRef]
- Alicata, J.E.; McCarthy, D.D. On the incidence and distribution of the rat lungworm Angiostrongylus cantonensis in the cook islands, with observations made in new zealand and western samoa. Can. J. Zool. 1964, 42, 605–611. [Google Scholar] [CrossRef]
- Kliks, M.M.; Palumbo, N.E. Eosinophilic meningitis beyond the Pacific Basin: The global dispersal of a peridomestic zoonosis caused by Angiostrongylus cantonensis, the nematode lungworm of rats. Soc. Sci. Med. 1992, 34, 199–212. [Google Scholar] [CrossRef]
- Alicata, J.E. The presence of Angiostrongylus cantonensis in islands of the Indian ocean and probable role of the giant African snail, achatina fulica, in dispersal of the parasite to the pacific islands. Can. J. Zool. 1966, 44, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Yousif, F.; Ibrahim, A. The first record of Angiostrongylus cantonensis from Egypt. Parasitol. Res. 1978, 56, 73–80. [Google Scholar] [CrossRef]
- Aguiar, P.H.; Morera, P.; Pascual, J. First record of Angiostrongylus cantonensis in Cuba. Am. J. Trop. Med. Hyg. 1981, 30, 963–965. [Google Scholar] [CrossRef]
- Vargas, M.; JD, G.P.; Malek, E.A. First record of Angiostrongylus cantonensis (Chen, 1935) (Nematoda: Metastrongylidae) in the Dominican Republic. Trop. Med. Parasitol. 1992, 43, 253–255. [Google Scholar] [PubMed]
- Campbell, B.G.; Little, M.D. The finding of Angiostrongylus cantonensis in rats in New Orleans. Am. J. Trop. Med. Hyg. 1988, 38, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Caldeira, R.L.; Mendonça, C.L.G.F.; Goveia, C.O.; Lenzi, H.L.; Graeff-Teixeira, C.; Lima, W.S.; Mota, E.M.; Pecora, I.L.; Medeiros, A.M.Z.D.; Carvalho, O.D.S. First record of molluscs naturally infected with Angiostrongylus cantonensis (Chen, 1935) (Nematoda: Metastrongylidae) in Brazil. Mem. Inst. Oswaldo Cruz 2007, 102, 887–889. [Google Scholar] [CrossRef]
- Foronda, P.; López-González, M.; Miquel, J.; Torres, J.; Segovia, M.; Abreu-Acosta, N.; Casanova, J.C.; Valladares, B.; Mas-Coma, S.; Bargues, M.D.; et al. Finding of Parastrongylus cantonensis (Chen, 1935) in Rattus rattus in Tenerife, Canary Islands (Spain). Acta Trop. 2010, 114, 123–127. [Google Scholar] [CrossRef]
- Paredes-Esquivel, C.; Sola, J.; Delgado-Serra, S.; Riera, M.P.; Negre, N.; Miranda, M.Á.; Jurado-Rivera, J.A. Angiostrongylus cantonensis in North African hedgehogs as vertebrate hosts, Mallorca, Spain, October 2018. Eurosurveillance 2019, 24, 1900489. [Google Scholar] [CrossRef] [Green Version]
- Mugisha, L.; Bwangamoi, O.; Cranfield, M. Angiostrongylus cantonensis and other parasites infections of rodents of Budongo Forest Reserve, Uganda. Afr. J. Anim. Biomed. Sci. 2012, 7, 91–95. [Google Scholar]
- Iwanowicz, D.D.; Sanders, L.R.; Schill, W.B.; Xayavong, M.V.; da Silva, A.J.; Qvarnstrom, Y.; Smith, T. Spread of the rat lungworm (Angiostrongylus cantonensis) in giant African land snails (Lissachatina fulica) in Florida, USA. J. Wildl. Dis. 2015, 51, 749–753. [Google Scholar] [CrossRef] [PubMed]
- Rizor, J.; Yanez, R.A.; Thaiwong, T.; Kiupel, M. Angiostrongylus cantonensis in a red ruffed lemur at a zoo, Louisiana, USA. Emerg. Infect. Dis. 2022, 28, 1058. [Google Scholar] [CrossRef]
- Dissanaike, A.S.; Cross, J.H. Ocular parastrongyliasis (=angiostrongyliasis): Probable first report of human infection from a patient in Ceylon (Sri Lanka). Parassitologia 2004, 46, 315–316. [Google Scholar] [PubMed]
- Limaye, L.S.; Pradhan, V.R.; Bhopale, M.K.; Renapurkar, D.M.; Sharma, K.D. Angiostrongylus cantonensis study of intermediate paratenic and definitive hosts in greater Bombay India. Helminthologia 1988, 25, 31–35. [Google Scholar]
- Limaye, L.S.; Bhopale, M.K.; Renapurkar, D.M.; Sharma, K.D. The distribution of Angiostrongylus cantonensis (Chen) in the central nervous system of laboratory rats. Folia Parasitol. 1983, 30, 281–284. [Google Scholar]
- Mahajan, R.; Almeida, A.; Sengupta, S.; Renapurkar, D. Seasonal intensity of Angiostrongylus cantonensis in the intermediate host, Laevicaulis alte. Int. J. Parasitol. 1992, 22, 669–671. [Google Scholar] [CrossRef] [PubMed]
- York, E.M.; Creecy, J.P.; Lord, W.D.; Caire, W. Geographic range expansion for rat lungworm in North America. Emerg. Infect. Dis. 2015, 21, 1234–1236. [Google Scholar] [CrossRef]
- Agrawal, V.C. Taxonomic Studies on Indian Muridae and Hystricidae (Mammalia: Rodentia); Rec. zool. Surv. India, Occasional Paper No. 180. i-viii; Director, Zoological Survey of India: Calcutta, India, 2000; pp. 1–186.
- Wilson, D.E.; Reeder, D.M. Mammal Species of the World: A Taxonomic and Geographic Reference, 3rd ed.; JHU Press: Baltimore, MA, USA, 2005; ISBN 0-8018-8221-4. [Google Scholar]
- Prakash, I.; Singh, P.; Nameer, P.; Ramesh, D.; Molur, S. South asian muroids. Mamm. S. Asia 2015, 2, 574–642. [Google Scholar]
- Wilson, D.E.; Lacher, T.E.; Mittermeier, R.A. Handbook of the Mammals of the World; Rodents II; Lynx Edicions: Barcelona, Spain, 2017; Volume 7. [Google Scholar]
- Dissanaike, A.S. The Proper Study of Mankind; Ceylon Association for the Advancement of Science: Colombo, Sri Lanka, 1968; pp. 115–142. [Google Scholar]
- Yong, H.S.; Eamsobhana, P. Definitive rodent hosts of the rat lungworm Angiostrongylus cantonensis. Raffles Bull. Zool. 2013, 29, 111–115. [Google Scholar]
- Adhikari, P.; Han, S.-H.; Kim, Y.-K.; Kim, T.-W.; Thapa, T.B.; Subedi, N.; Kunwar, A.; Banjade, M.; Oh, H.-S. New record of the Oriental house rat, Rattus tanezumi, in Nepal inferred from mitochondrial cytochrome B gene sequences. Mitochondrial DNA Part B 2018, 3, 386–390. [Google Scholar] [CrossRef] [Green Version]
- Aplin, K.P.; Brown, P.R.; Jacob, J.; Krebs, C.J.; Singleton, G.R. Field Methods for Rodent Studies in Asia and the Indo-Pacific; Australian Centre for International Agricultural Research: Melbourne Australia, 2003; ISBN 1863203931.
- Bentley, E.; Bathard, A. The rats of Addu Atoll, Maldive Islands. Ann. Mag. Nat. Hist. 1959, 2, 365–368. [Google Scholar] [CrossRef]
- Cowie, R.H. Biology, systematics, life cycle, and distribution of Angiostrongylus cantonensis, the cause of rat lungworm disease. Hawaii J. Med. Public Health 2013, 72, 6. [Google Scholar] [PubMed]
- Lv, S.; Zhang, Y.; Liu, H.-X.; Hu, L.; Yang, K.; Steinmann, P.; Chen, Z.; Wang, L.-Y.; Utzinger, J.; Zhou, X.-N. Invasive snails and an emerging infectious disease: Results from the first national survey on Angiostrongylus cantonensis in China. PLoS Negl. Trop. Dis. 2009, 3, e368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, C.S.; Merritt, J.W. Studies on Angiostrongylus cantonensis in molluscan intermediate hosts. J. Parasitol. 1967, 53, 382. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.; Mukhopadhayay, A. Freshwater molluscs of India: An insight of into their diversity, distribution and conservation. In Aquatic Ecosystem: Biodiversity, Ecology and Conservation; Springer: New Delhi, India, 2015; pp. 163–195. [Google Scholar] [CrossRef]
- Sen, S.; Ravikanth, G.; Aravind, N. Land snails (Mollusca: Gastropoda) of India: Status, threats and conservation strategies. J. Threat. Taxa 2012, 4, 3029–3037. [Google Scholar] [CrossRef] [Green Version]
- Boominathan, M.; Ravikumar, G.; Chandran, M.D.S.; Ramachandra, T.V. Mangrove associated molluscs of India. In Proceedings of the National Conference on Conservation and Management of Wetland Ecosystems, Kerala, India, 6–9 November 2012; Volume 7, pp. 1–11. [Google Scholar]
- Ratnapala, R. Land snails: Distribution and notes on ecology. In Ecology and Biogeography in Sri Lanka; Springer: Dordrecht, The Netherlands, 1984; pp. 391–411. [Google Scholar] [CrossRef]
- Strong, E.E.; Gargominy, O.; Ponder, W.; Bouchet, P. Global diversity of gastropods (Gastropoda; Mollusca) in freshwater. Freshw. Anim. Divers. Assess. 2008, 198, 149–166. [Google Scholar] [CrossRef]
- Mavinkurve, R.G.; Shanbhag, S.P.; Madhyastha, N. Non-marine molluscs of Western Ghats: A status review. Zoos’ Print J. 2004, 19, 1708–1711. [Google Scholar] [CrossRef]
- Thiengo, S.; Maldonado, A.; Mota, E.; Torres, E.; Caldeira, R.; Carvalho, O.; Oliveira, A.; Simões, R.; Fernandez, M.; Lanfredi, R. The giant African snail Achatina fulica as natural intermediate host of Angiostrongylus cantonensis in Pernambuco, northeast Brazil. Acta Trop. 2010, 115, 194–199. [Google Scholar] [CrossRef]
- Wang, Q.-P.; Chen, X.-G.; Lun, Z.-R. Invasive fresh water snail, China. Emerg. Infect. Dis. 2007, 13, 1119–1120. [Google Scholar] [CrossRef]
- Cowie, R.H. Angiostrongylus cantonensis: Agent of a sometimes fatal globally emerging infectious disease (rat lungworm disease). ACS Chem. Neurosci. 2017, 8, 2102–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eamsobhana, P.; Yong, H.S.; Prasartvit, A.; Wanachiwanawin, D.; Boonyong, S. Geographical distribution and incidence of Angiostrongylus lungworms (Nematoda: Angiostrongylidae) and their rodent hosts in Thailand. Trop. Biomed. 2016, 33, 35–44. [Google Scholar] [PubMed]
- Hamilton, L.J.; Tagami, Y.; Kaluna, L.; Jacob, J.; Jarvi, S.I.; Follett, P. Demographics of the semi-slug Parmarion martensi, an intermediate host for Angiostrongylus cantonensis in Hawai‘i, during laboratory rearing. Parasitology 2021, 148, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.R.; Wong, T.M.; Curry, P.A.; Yeung, N.W.; Hayes, K.A.; Cowie, R.H. Modelling the distribution in Hawaii of Angiostrongylus cantonensis (rat lungworm) in its gastropod hosts. Parasitology 2019, 146, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Tesana, S.; Srisawangwong, T.; Sithithaworn, P.; Laha, T.; Andrews, R. Prevalence and intensity of infection with third stage larvae of Angiostrongylus cantonensis in mollusks from Northeast Thailand. Am. J. Trop. Med. Hyg. 2009, 80, 983–987. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, W.; Yao, J.; Yang, M.; Guo, Y.; Deng, Z.; Mao, Q.; Li, S.; Duan, L. Comparative proteomics suggests the mode of action of a novel molluscicide against the invasive apple snail Pomacea canaliculata, intermediate host of Angiostrongylus cantonensis. Mol. Biochem. Parasitol. 2022, 247, 111431. [Google Scholar] [CrossRef]
- Yang, T.-B.; Wu, Z.-D.; Lun, Z.-R. The apple snail Pomacea canaliculata, a novel vector of the rat lungworm, Angiostrongylus cantonensis: Its introduction, spread, and control in China. Hawaii J. Med. Public Health 2013, 72, 23. [Google Scholar]
- Baloch, W.A.; Memon, U.N.; Burdi, G.H.; Soomro, A.N.; Tunio, G.R.; Khatian, A.A. Invasion of channelled apple snail Pomacea canaliculata, Lamarck (Gastropoda: Ampullariidae) in Haleji Lake, Pakistan. Sindh Univ. Res. J. SURJ (Sci. Ser.) 2012, 44, 263–266. [Google Scholar]
- Joshi, R.C. Problems with the management of the golden apple snail Pomacea canaliculata: An important exotic pest of rice in Asia. In Proceedings of the Area-Wide Control of Insect Pests: From Research to Field Implementation; Springer: Berlin/Heidelberg, Germany, 2007; pp. 257–264. [Google Scholar]
- Rekha Sarma, R.; Munsi, M.; Neelavara Ananthram, A. Effect of climate change on invasion risk of giant African snail (Achatina fulica Férussac, 1821: Achatinidae) in India. PLoS ONE 2015, 10, e0143724. [Google Scholar] [CrossRef] [Green Version]
- Saha, C.; Pramanik, S.; Chakraborty, J.; Parveen, S.; Aditya, G. Abundance and body size of the invasive snail Physa acuta occurring in Burdwan, West Bengal, India. J. Entomol. Zool. Stud. 2016, 4, 490–497. [Google Scholar]
- Saha, C.; Parveen, S.; Chakraborty, J.; Pramanik, S.; Aditya, G. Life table estimates of the invasive snail Physa acuta Draparnaud, 1805, occurring in India. Ekologia 2017, 36, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Paul, P.; Aditya, G. Invasion of the freshwater snail Physella acuta (Draparnaud, 1805) in selected ponds of North Dinajpur, India. J. Environ. Biol. 2021, 42, 577–581. [Google Scholar] [CrossRef]
- Raut, S.K.; Mandal, R.N. Natural history of the garden slug Laevicaulis alte. J. Bengal Nat. Hist. Soc. 1984, 3, 104–105. [Google Scholar]
- Thakuri, B.; Acharya, B.K.; Sharma, G. Population density and damage of invasive giant African snail Achatina fulica in organic farm in east Sikkim, India. Indian J. Ecol. 2019, 46, 631–635. [Google Scholar]
- Sridhar, V.; Vinesh, L.S.; Jayashankar, M. Mapping the potential distribution of Achatina fulica (Bowdich) (Stylommatophora: Achatinidae) in India using CLIMEX, a bioclimatic software. Pest Manag. Hortic. Ecosyst. 2014, 20, 14–21. [Google Scholar]
- Husain, A.; Husain, H.J.; Hasan, W.; Kendra, J.K.V. New records of tropical leather-leaf slug Laevicaulis alte (Ferussac, 1822) from Dehra Dun (Uttarakhand) and Jamshedpur (Jharkhand), India. Int. J. Agric. Appl. Sci. 2021, 2, 145–150. [Google Scholar] [CrossRef]
- Baghele, M.; Mishra, S.; Meyer-Rochow, V.B.; Jung, C.; Ghosh, S. Utilization of snails as food and therapeutic agents by Baiga tribals of Baihar tehsil of Balaghat District, Madhya Pradesh, India. Int. J. Indust. Entomol. 2021, 43, 78–84. [Google Scholar]
- Dhiman, V.; Pant, D. Human health and snails. J Immunoassay Immunochem. 2021, 42, 211–235. [Google Scholar] [CrossRef] [PubMed]
- Prabhakar, A.K.; Roy, S. Ethno-medicinal uses of some shell fishes by people of Kosi river basin of North-Bihar, India. Stud. Ethno-Med. 2009, 3, 1–4. [Google Scholar] [CrossRef]
- Sarkar, T.; Debnath, S.; Das, B.K.; Das, M. Edible fresh water molluscs diversity in the different water bodies of Gangarampur Block, Dakshin Dinajpur, West Bengal. Eco. Env. Cons. 2021, 27, S293–S296. [Google Scholar]
- Baby, R.L.; Hasan, I.; Kabir, K.A.; Naser, M.N. Nutrient analysis of some commercially important molluscs of Bangladesh. J. Sci. Res. 2010, 2, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Rabha, H.P.; Mazumdar, M.; Baruah, U.K. Indigenous technical knowledge on ethnic dishes of snail in Goalpara district of India. Sch. Acad. J. Biosci. 2014, 2, 307–317. [Google Scholar]
- Borkakati, R.N.; Gogoi, R.; Borah, B.K. Snail: From present perspective to the history of Assam. Asian Agrihist. 2009, 13, 227–234. [Google Scholar]
- Ghosh, S.; Jung, C.; Meyer-Rochow, V.B. Snail farming: An Indian perspective of a potential tool for food security. Ann. Aquac. Res. 2016, 3, 1024. [Google Scholar]
- Bagde, N.; Jain, S. Study of traditional man-animal relationship in Chhindwara district of Madhya Pradesh, India. J. Glob. Biosci. 2015, 4, 1456–1463. [Google Scholar]
- Tsai, H.-C.; Lin, H.-H.; Wann, S.-R.; Liu, Y.-C.; Huang, C.-K.; Lai, P.-H.; Yen, M.-Y.; Kunin, C.M.; Ger, L.-P.; Lee, S.S.-J.; et al. Eosinophilic meningitis caused by Angiostrongylus cantonensis associated with eating raw snails: Correlation of brain magneticresonance imaging scans with clinical findings. Am. J. Trop. Med. Hyg. 2003, 68, 281–285. [Google Scholar] [CrossRef]
- Alicata, J.E. Effect of freezing and boiling on the infectivity of third-stage larvae of Angiostrongylus cantonensis present in and snails and freshwater prawns. J. Parasitol. 1967, 53, 1064. [Google Scholar] [CrossRef] [PubMed]
- Crook, J.R.; Fulton, S.E.; Supanwong, K. The infectivity of third stage Angiostrongylus cantonensis larvae shed from drowned Achatina fulica snails and the effect of chemical agents on infectivity. Trans. R. Soc. Trop. Med. Hyg. 1971, 65, 602–605. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Kumar, M.U.; Kumari, A. Freshwater shellfish, Pila globosa (Gastropoda) favourable endeavour for rural nutrition. Agric. Lett. 2021, 2, 32–35. [Google Scholar]
- Radomyos, P.; Tungtrongchitr, A.; Praewanich, R. Experimental infection of yellow tree monitor (Varanus bengalensis) and related species with Angiostrongylus cantonensis. Southeast Asian J. Trop. Med. Public Health 1992, 23, 167–168. [Google Scholar] [PubMed]
- Böhme, W. Checklist of the living monitor lizards of the world (Family Varanidae). Zool. Verh. 2003, 341, 4–43. [Google Scholar]
- Pianka, E.; King, D. Varanoid Lizards of the World; Indiana University Press: Bloomington, IN, USA, 2004; ISBN 0253013690. [Google Scholar]
- Aengals, R.; Kumar, V.S.; Palot, M.J.; Ganesh, S.R. A Checklist of Reptiles of India; Zoological Survey of India: Kolkata, India, 2018; p. 35.
- Auffenberg, W.; Rahman, H.; Iffat, F.; Perveen, Z. A study of Varanus flavescens (Hardwicke & Gray) (Sauria: Varanidae). J. Bombay Nat. Hist. Soc. 1989, 86, 286–307. [Google Scholar]
- Wikramanayake, E.D.; Dryden, G.L. Thermal ecology of habitat and microhabitat use by sympatric Varanus bengalensis and V. salvator in Sri Lanka. Copeia 1993, 709–714. [Google Scholar] [CrossRef]
- Radomyos, P.; Tungtrongchitr, A.; Praewanich, R.; Khewwatchan, P.; Kantangkul, T.; Junlananto, P.; Ayudhya, S.I. Occurrence of the infective stage of Angiostrongylus cantonensis in the yellow tree monitor (Varanus bengalensis) in five Provinces of Thailand. Southeast Asian J. Trop. Med. Public Health 1994, 25, 498–500. [Google Scholar]
- Anettová, L.; Izquierdo-Rodriguez, E.; Foronda, P.; Baláž, V.; Novotný, L.; Modrý, D. Endemic lizard Gallotia galloti is a paratenic host of invasive Angiostrongylus cantonensis in Tenerife, Spain. Parasitology 2022, 149, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Darasavath, C.; Chang, K.; Vilay, V.; Sengduangphachanh, A.; Adsamouth, A.; Vongsouvath, M.; Keolouangkhot, V.; Robinson, M.T. Cluster of angiostrongyliasis cases following consumption of raw monitor lizard in the Lao People’s Democratic Republic and review of the literature. Trop. Med. Infect. Dis. 2021, 6, 107. [Google Scholar] [CrossRef] [PubMed]
- Ash, L.R. The Occurrence of Angiostrongylus cantonensis in frogs of New Caledonia with observations on paratenic hosts of metastrongyles. J. Parasitol. 1968, 54, 432. [Google Scholar] [CrossRef]
- Chellappandian, M.; Pandikumar, P.; Mutheeswaran, S.; Paulraj, M.G.; Prabakaran, S.; Duraipandiyan, V.; Ignacimuthu, S.; Al-Dhabi, N. Documentation and quantitative analysis of local ethnozoological knowledge among traditional healers of Theni district, Tamil Nadu, India. J. Ethnopharmacol. 2014, 154, 116–130. [Google Scholar] [CrossRef]
- Jugli, S.; Chakravorty, J.; Meyer-Rochow, V.B. Zootherapeutic uses of animals and their parts: An important element of the traditional knowledge of the Tangsa and Wancho of Eastern Arunachal Pradesh, North-East India. Environ. Dev. Sustain. 2020, 22, 4699–4734. [Google Scholar] [CrossRef] [Green Version]
- Chakravorty, J.; Meyer-Rochow, V.B.; Ghosh, S. Vertebrates used for medicinal purposes by members of the Nyishi and Galo tribes in Arunachal Pradesh (North-East India). J. Ethnobiol. Ethnomed. 2011, 7, 13. [Google Scholar] [CrossRef] [Green Version]
- Das, K.S.; Choudhury, S.; Nonglait, K.C.L. Zootherapy among the ethnic groups of north eastern region of india-a critical review. J. Crit. Rev. 2017, 4, 1–9. [Google Scholar] [CrossRef]
- Mahawar, M.M.; Jaroli, D.P. Animals and their products utilized as medicines by the inhabitants surrounding the Ranthambhore National Park, India. J. Ethnobiol. Ethnomed. 2006, 2, 46. [Google Scholar] [CrossRef] [Green Version]
- Verma, A.K.; Prasad, S.B.; Rongpi, T.; Arjun, J. Traditional healing with animals (zootherapy) by the major ethnic group of Karbi Anglong district of Assam, India. Int. J. Pharm. Pharm. Sci. 2014, 6, 593–600. [Google Scholar]
- Borah, M.P.; Prasad, S.B. Ethnozoological study of animals based medicine used by traditional healers and indigenous inhabitants in the adjoining areas of Gibbon Wildlife Sanctuary, Assam, India. J. Ethnobiol. Ethnomed. 2017, 13, 39. [Google Scholar] [CrossRef] [Green Version]
- Betlu, A.L.S. Indigenous knowledge of zootherapeutic use among the Biate tribe of Dima Hasao District, Assam, Northeastern India. J. Ethnobiol. Ethnomed. 2013, 9, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oku, Y.; Katakura, K.; Kamiya, M. Tadpole of the clawed frog, Xenopus Laevis, as an experimental intermediate host of Angiostrongylus cantonensis. Am. J. Trop. Med. Hyg. 1980, 29, 316–318. [Google Scholar] [CrossRef] [PubMed]
- Niebuhr, C.N.; Jarvi, S.I.; Kaluna, L.; Fischer, B.L.T.; Deane, A.R.; Leinbach, I.L.; Siers, S.R. Occurrence of rat lungworm (Angiostrongylus cantonensis) in invasive coqui frogs (Eleutherodactylus coqui) and other hosts in Hawaii, USA. J. Wildl. Dis. 2020, 56, 203. [Google Scholar] [CrossRef] [PubMed]
- Asato, R.; Sato, Y.; Otsuru, M. The occurrence of Angiostrongylus cantonensis in toads and frogs in Okinawa Prefecture, Japan. Jpn J. Parasitol. 1978, 27, 1–8. [Google Scholar]
- Mahawar, M.M.; Jaroli, D. Traditional zootherapeutic studies in India: A review. J. Ethnobiol. Ethnomed. 2008, 4, 17. [Google Scholar] [CrossRef] [Green Version]
- Shah, K.B.; Gire, M.K. Some Amphibians and Their Local Uses in Arun Basin. J. Nat. Hist. Museum. 1992, 13, 9–17. [Google Scholar]
- Pradhan, S.; Mishra, D.; Sahu, K.R. Herpetofauna used as traditional medicine by tribes of Gandhamardan Hills Range, Western Orissa, India. Int. J. Res. Zool. 2014, 4, 32–35. [Google Scholar]
- Pandian, T.J.; Marian, M.P. Production and utilization of frogs: An ecological view. Proc. Anim. Sci. 1986, 95, 289–301. [Google Scholar] [CrossRef]
- Kawlni, L.; Lalronunga, S.; Manral, U.; Das, A. Uchang-Sa: A report on frog consumption around Dampa Tiger Reserve, Mizoram, Northeast India. Hamadryad 2022, 39, 30–36. [Google Scholar]
- Talukdar, S.; Sengupta, S. Edible frog species of Nagaland. J. Environ. Biol. 2020, 41, 927–930. [Google Scholar] [CrossRef]
- Talukdar, S.; Sengupta, S.; Konyak, M.; Rizwan, M. Cooking methods and medicinal uses of frog species among the Naga tribes in Dimapur. World News Nat. Sci. 2020, 30, 144–149. [Google Scholar]
- Thomas, A.; Biju, S.D. Tadpole consumption is a direct threat to the endangered purple frog, Nasikabatrachus Sahyadrensis. Salamandra 2015, 51, 252–258. [Google Scholar]
- Yong, H.-S.; Eamsobhana, P.; Song, S.-L.; Prasartvit, A.; Lim, P.-E. Molecular phylogeography of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae) and genetic relationships with congeners using cytochrome b gene marker. Acta Trop. 2015, 148, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Yong, H.-S.; Song, S.-L.; Eamsobhana, P.; Goh, S.-Y.; Lim, P.-E. Complete mitochondrial genome reveals genetic diversity of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae). Acta Trop. 2015, 152, 157–164. [Google Scholar] [CrossRef]
- Vitta, A.; Srisongcram, N.; Thiproaj, J.; Wongma, A.; Polsut, W.; Fukruksa, C.; Yimthin, T.; Mangkit, B.; Thanwisai, A.; Dekumyoy, P. Phylogeny of Angiostrongylus cantonensis in Thailand based on cytochrome c oxidase subunit I gene sequence. Southeast Asian J. Trop. Med. Public Health 2016, 47, 377–386. [Google Scholar]
- Dumidae, A.; Subkrasae, C.; Ardpairin, J.; Thanwisai, A.; Vitta, A. Genetic analysis of a 66-kDa protein-encoding gene of Angiostrongylus cantonensis and Angiostrongylus malaysiensis. Parasitology 2023, 150, 98–114. [Google Scholar] [CrossRef]
- Rodpai, R.; Intapan, P.M.; Thanchomnang, T.; Sanpool, O.; Sadaow, L.; Laymanivong, S.; Aung, W.P.; Phosuk, I.; Laummaunwai, P.; Maleewong, W. Angiostrongylus cantonensis and A. malaysiensis broadly overlap in Thailand, Lao PDR, Cambodia and Myanmar: A molecular survey of larvae in land snails. PLoS ONE 2016, 11, e0161128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, A.H.E.; Chaisiri, K.; Dusitsittipon, S.; Jakkul, W.; Charoennitiwat, V.; Komalamisra, C.; Thaenkham, U. Mitochondrial ribosomal genes as novel genetic markers for discrimination of closely related species in the Angiostrongylus cantonensis lineage. Acta Trop. 2020, 211, 105645. [Google Scholar] [CrossRef] [PubMed]
- Eamsobhana, P.; Lim, P.; Yong, H. Phylogenetics and systematics of Angiostrongylus lungworms and related taxa (Nematoda: Metastrongyloidea) inferred from the nuclear small subunit (SSU) ribosomal DNA sequences. J. Helminthol. 2014, 89, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Eamsobhana, P.; Song, S.-L.; Yong, H.-S.; Prasartvit, A.; Boonyong, S.; Tungtrongchitr, A. Cytochrome c oxidase subunit I haplotype diversity of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae). Acta Trop. 2017, 171, 141–145. [Google Scholar] [CrossRef]
- Eamsobhana, P.; Yong, H.-S.; Song, S.-L.; Gan, X.-X.; Prasartvit, A.; Tungtrongchitr, A. Molecular phylogeography and genetic diversity of Angiostrongylus cantonensis and A. malaysiensis (Nematoda: Angiostrongylidae) based on 66-kDa protein gene. Parasitol. Int. 2019, 68, 24–30. [Google Scholar] [CrossRef]
- Mishra, N.; Panda, T. Zootherapeutical Uses of Animal Diversity in Coastal District of Orissa, India. Br. J. Pharmacol. Toxicol. 2011, 2, 154–158. [Google Scholar]
- Joshi, N.; Pandey, S. Meat Demand-Snailed It: A Comprehensive Review on Snail Rearing, to Meet the Meat Demand in Future India. J. Entomol. Zool. Stud 2019, 7, 396–400. [Google Scholar]
- MESHRAM, L.; HUSAIN, N. Use of Animals as Traditional Medicine in Chhattisgarh, India. Indian J. Sci. Res 2017, 12, 26–29. [Google Scholar]
- Raja, L.; Matheswaran, P.; Anbalagan, M.; Sureshkumar, V.; Ganesan, D.; Gani, S.B. Ethnozoological Study of Animal-Based Products Practices among the Tribal Inhabitants in Kolli Hills Namakkal District, Tamil Nadu, India. World J. Pharm. Pharm. Sci. 2018, 7, 785–797. [Google Scholar]
- Debnath, C.; Sahoo, L.; Haldar, A.; Datta, M.; Yadav, G.S.; Singha, A.; Bhattacharjee, J. Proximate and Mineral Composition of Freshwater Snails of Tripura, North-East India. Fish. Technol. 2016, 53, 307–312. [Google Scholar]
- Chinlampianga, M.; Singh, R.K.; Shukla, A.C. Ethnozoological Diversity of Northeast India: Empirical Learning with Traditional Knowledge Holders of Mizoram and Arunachal Pradesh; NISCAIR-CSIR: Delhi, India, 2013. [Google Scholar]
- Singh, R.K.; Kumar, A.; Singh, A.; Singhal, P. Evidence That Cultural Food Practices of Adi Women in Arunachal Pradesh, India, Improve Social-Ecological Resilience: Insights for Sustainable Development Goals. Ecol. Process. 2020, 9, 1–19. [Google Scholar] [CrossRef]
- Jadhav, A.; Das, N.K.; Aravind, N.A. Edible Freshwater Molluscs from Northeast India. Tentacle 2020, 28, 1–4. [Google Scholar]
- Bhattacharya, S.; Chakraborty, M.; Bose, M.; Mukherjee, D.; Roychoudhury, A.; Dhar, P.; Mishra, R. Indian Freshwater Edible Snail Bellamya Bengalensis Lipid Extract Prevents T Cell Mediated Hypersensitivity and Inhibits LPS Induced Macrophage Activation. J. Ethnopharmacol. 2014, 157, 320–329. [Google Scholar] [CrossRef] [PubMed]
- Gomes, A.; Sarkar, A.; Ghosh, S.; Saha, K.; Saha, P.P. Edible Fresh Water Snail Viviparous Bengalensis Purified Flesh Protein VB-P4 Induced Toxicities and Its Protection by Heat Treatment. J. Toxins 2015, 2, 3. [Google Scholar]
- Dhakal, P.; Chettri, B.; Lepcha, S.; Acharya, B.K. Rich yet Undocumented Ethnozoological Practices of Socio-Culturally Diverse Indigenous Communities of Sikkim Himalaya, India. J. Ethnopharmacol. 2020, 249, 112386. [Google Scholar] [CrossRef]
- Patel, R.; Pal, M.C. Exploring Ethnozoological Observations Among The Tribal Inhabitants In ‘Bajag’ Forest Range Of Dindori District Of Central India. Int. J. Aquat. Sci. 2021, 12, 4513–4538. [Google Scholar]
- Vyas, N.; Mahawar, M.M.; Jaroli, D.P. Traditional Medicines Derived from Domestic Animals Used by Rebari Community of Rajasthan, India. Nature 1970, 7, 129–138. [Google Scholar] [CrossRef] [Green Version]
- Solavan, A.; Paulmurugan, R.; Wilsanand, V.; Sing, A.J. Traditional Therapeutic Uses of Animals among Tribal Population of Tamil Nadu; CSIR: Lucknow, India, 2004. [Google Scholar]
- Haq, S.M.; Calixto, E.S.; Yaqoob, U.; Ahmed, R.; Mahmoud, A.H.; Bussmann, R.W.; Mohammed, O.B.; Ahmad, K.; Abbasi, A.M. Traditional Usage of Wild Fauna among the Local Inhabitants of Ladakh, Trans-Himalayan Region. Animals 2020, 10, 2317. [Google Scholar] [CrossRef]
- Imkongwapang, R.; Jyrwa, D.B.; Lal, P.; Tandon, V. A Checklist of Helminth Parasite Fauna in Anuran Amphibia (Frogs) of Nagaland, Northeast India. J. Parasit. Dis. 2014, 38, 85–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, M.; Haq, S.M.; Amjad, M.S.; Ahmad, R.; Bussmann, R.W.; de la Lastra, J.M.P. Invertebrates and Herptiles for Livelihoods—Ethnozoological Use among Different Ethnic Communities in Jammu and Kashmir (Indian Himalayas). Front. Pharmacol. 2023, 13, 1043155. [Google Scholar] [CrossRef] [PubMed]
- Narzary, J.; Bordoloi, S. Ethnozoological Practices on Frogs of Bodo Tribe from Kokrajhar District, Assam, India. Am. J. Ethnomedicine 2014, 1, 368–370. [Google Scholar]
- Vijayakumar, S.; Yabesh, J.E.M.; Prabhu, S.; Ayyanar, M.; Damodaran, R. Ethnozoological Study of Animals Used by Traditional Healers in Silent Valley of Kerala, India. J. Ethnopharmacol. 2015, 162, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.; Sreeedharan, S.; George, S.; Antony, M.M. The Complete Mitochondrial Genome of an Endemic Cichlid Etroplus Canarensis from Western Ghats, India (Perciformes: Cichlidae) and Molecular Phylogenetic Analysis. Mol. Biol. Rep. 2022, 49, 3033–3044. [Google Scholar] [CrossRef]
- Esakkimuthu, S.; Sylvester Darvin, S.; Mutheeswaran, S.; Gabriel Paulraj, M.; Pandikumar, P.; Ignacimuthu, S.; Al-Dhabi, N.A. A Study on Food-Medicine Continuum among the Non-Institutionally Trained Siddha Practitioners of Tiruvallur District, Tamil Nadu, India. J. Ethnobiol. Ethnomed. 2018, 14, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar Kiran, S.; Anoop, V.S.; Sivakumar, K.C.; Dinesh, R.; Mano, J.P.; Kaushik, D.; Sanil, G. An Additional Record of Fejervarya Manoharani Garg and Biju from the Western Ghats with a Description of Its Complete Mitochondrial Genome. Zootaxa 2017, 4277, 491–502. [Google Scholar] [CrossRef]
- Anoop, V.S.; George, S. Population Genetic Structure and Evolutionary Demographic Patterns of Phrynoderma Karaavali, an Edible Frog Species of Kerala, India. J. Genet. 2023, 102, 8. [Google Scholar] [CrossRef]
- Chettri, B.; Bhupathy, S.; Acharya, B.K. An Overview of the Herpetofauna of Sikkim with Emphasis on the Elevational Distribution Pattern and Threats and Conservation Issues. In Biodiversity of Sikkim: Exploring and Conserving a Global Hotspot; Information and Public Relations Department, Government of Sikkim: Gangtok, India, 2011; pp. 233–254. [Google Scholar]
- Jaroli, D.P.; Mahawar, M.M.; Vyas, N. An Ethnozoological Study in the Adjoining Areas of Mount Abu Wildlife Sanctuary, India. J. Ethnobiol. Ethnomed. 2010, 6, 6. [Google Scholar] [CrossRef] [Green Version]



| Country | State | Bandicota indica (Rodentia: Muridae) | Rattus norvegicus (Rodentia: Muridae) | Laevicaulis alte (Gastropoda: Veronicellidae) | Macrochlamys indica (Gastropoda: Ariophantidae) | Reference |
|---|---|---|---|---|---|---|
| India | Kerala | + | [41] | |||
| India | Maharashtra | + | + | + | [95,96] | |
| India | Tamil nandu | + | [10,83] | |||
| Sri Lanka | Ceylon | + | + | [10,83] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pandian, D.; Najer, T.; Modrý, D. An Overview of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae), an Emerging Cause of Human Angiostrongylosis on the Indian Subcontinent. Pathogens 2023, 12, 851. https://doi.org/10.3390/pathogens12060851
Pandian D, Najer T, Modrý D. An Overview of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae), an Emerging Cause of Human Angiostrongylosis on the Indian Subcontinent. Pathogens. 2023; 12(6):851. https://doi.org/10.3390/pathogens12060851
Chicago/Turabian StylePandian, Divakaran, Tomáš Najer, and David Modrý. 2023. "An Overview of Angiostrongylus cantonensis (Nematoda: Angiostrongylidae), an Emerging Cause of Human Angiostrongylosis on the Indian Subcontinent" Pathogens 12, no. 6: 851. https://doi.org/10.3390/pathogens12060851