
Cacopsylla fuscicella Sp. Nov. (Hemiptera, Psyllidae), a New Loquat Pest in China †


1
Department of Entomology MOA Key Lab of Pest Monitoring and Green Management, College of Plant Protection, China Agricultural University, Beijing 100193, China
2
Zhaotong Green Food Development Center, Zhaotong, Yunnan 657000, China
*
Author to whom correspondence should be addressed.
†
LSID urn:lsid:zoobank.org:pub:FBC10EFF-3B4C-4424-B782-B7B3AF0FBDA3.
Insects 2023, 14(5), 414; https://doi.org/10.3390/insects14050414
Submission received: 14 March 2023
/
Revised: 22 April 2023
/
Accepted: 23 April 2023
/
Published: 26 April 2023
(This article belongs to the Section Insect Systematics, Phylogeny and Evolution)
Abstract
:Simple Summary
Cacopsylla, the most species-rich genus of Psyllidae, has around 500 known species worldwide. These insects are known for being host-specific, and some of them are regarded as important pests. Here, we described a new species, Cacopsylla fuscicella sp. nov., which develops on loquats. In addition, we sequenced its mitochondrial genome and used these data to construct a phylogenetic tree of Psyllidae, while also calculating the interspecific genetic distances. Using integrative taxonomy methods, we confirmed that the new species, Cacopsylla fuscicella sp. nov., should be grouped within genus Cacopsylla.
Abstract
Psylloidea, commonly known as jumping plant-lice, have a phloem-sucking habit and strong specificity to their host plants. Within the Psyllidae family, the genus Cacopsylla Ossiannilsson, 1970 stands out as being the most diverse, with three species feeding on the genus Eriobotrya Lindl. In this study, a new psyllid species, Cacopsylla fuscicella sp. nov., was described from China. It is a pest of Eriobotrya japonica (Thunb.) Lindl. and has been cultivated as a commercial fruit tree for years. Illustrations of habitus, morphological structures, and its damage to loquat were also provided. The complete mitochondrial genome of C. fuscicella sp. nov. was sequenced and annotated. The phylogenetic tree inferred from the maximum likelihood analyses confirmed C. fuscicella sp. nov. as a member of the genus Cacopsylla. Genetic distances were generated to assess comparative genetic distances within Psyllinae.
1. Introduction
The Psylloidea, commonly known as jumping plant lice, are a group of phytophagous insects with a phloem-sucking habit, around 1–6 mm in size. Some species are important pests, damaging crops and trees and even vectoring plant diseases [1]. Generally, psyllids are host-specific species. The range of host plants for a certain psyllid species is usually one genus or several genera belonging to the same family of plant. Currently, Cacopsylla Ossiannilsson, 1970 is the most diverse genus in the family Psyllidae, with nearly 500 known species distributed worldwide [2]. Of these, around 400 species have been recorded to occur in China [3]. This genus has been considered polyphyletic by multiple authors, with different species feeding on distinct plant groups, putatively forming many species groups. Consequently, numerous species remain disputed or ungrouped [4,5]. A considerable number of Cacopsylla species are known to feed on various Rosaceae plants. These species have historically been assigned to different subgenera [2], but these subgenera are likely not monophyletic and need further clarification. For example, different members feeding on Pyrus spp. from western and eastern Palaearctic regions have been considered to be distinctly related. This implies at least two independent origins for these species, which may have evolved as a result of separately occurring host shifting events [6,7]. These thus leave the pattern of evolution and taxonomy of Cacopsylla problematic and intriguing.
Molecular methods have been used to help elucidate problems of taxonomy and evolutionary patterns in insects. Integrative taxonomy is widely regarded as a promising method for resolving taxonomic issues. For example, Cho et al. [8] analyzed DNA barcodes of 11 pear psyllid species to confirm synonymies and correct misidentifications. Mitochondrial genes are commonly used in phylogenetic analyses. Percy et al. [4] used more than 75 complete mitochondrial genomes of Psyllidae taxa in their phylogenetic study, with eight species being annotated and uploaded to GenBank. Similarly, 13 protein coding genes (PCGs) of two and three species of Cacopsylla were used to build phylogenetic trees of Psyllidae and Psylloidea, respectively [9,10]. Currently, 12 annotated mitochondrial genomes (mitogenomes) of Psyllidae species are available in GenBank [4,9,10,11,12,13].
The Cacopsylla species, which are associated with Rosaceae, also include several significant agricultural pests. The Chinese pear psyllid, Cacopsylla chinensis (Yang and Li, 1981), for example, has long been a major pest in pear orchards in China [14]; while the West Palearctic species, Cacopsylla pyricola (Förster, 1848), is an important pear pest in Europe and also known as the vector of the fire blight [15,16]. Similar concerns are continuously arising with the discovery of more Cacopsylla species associated with this fruit-rich family; some species have been treated as local endemic pests [7,17].
A rather small group of Cacopsylla including three species dealt with in this research feeds on Eriobotrya Lindl., a genus of the angiosperm family Rosaceae. The three species are Cacopsylla eriobotryacola (Yang, 1984), Cacopsylla eriobotryae (Yang, 1984), and Cacopsylla biwa (Inoue, 2013). C. eriobotryacola and C. eriobotryae are associated with the species Eriobotrya deflexa (Hemsl.) Nakai; and the other one, C. biwa, is the pest of E. japonica [18,19]. Eriobotrya japonica (Thunb.) Lindl., also known as loquat or biwa, is cultivated for its fruit and has become the most important commercial fruit tree among this genus. It is being increasingly cultivated in Asia, especially in China, where it even became the major income source for the local fruit growers. The new species described here was found in loquat orchards in Zhaotong City, Yunnan Province, where they occur abundantly and cause considerable damage.
In the present study, we describe a new Cacopsylla species feeding on loquats, providing characters of both adults and fifth-instar immatures. We also sequenced its mitochondrial genome and compare genetic distances within Psyllinae. A phylogenetic tree using 13 PCGs and 2 rRNA genes of Psyllidae was constructed to discuss the systematic position of Cacopsylla fuscicella sp. nov. relative to other species of Cacopsylla and Psyllidae sensu stricto.
2. Materials and Methods
2.1. Morphological Study
Specimens were boiled in 10% potassium hydroxide (KOH) (Sinopharm Chemical Reagent Co., Ltd., Beijing, China) solution for 10 min, naturally cooled down, washed in distilled water, and mounted on a slide in glycerin (Sinopharm Chemical Reagent Co., Ltd., Beijing, China) for further examination. Measurements were taken from slide-mounted specimens. For adults: BL = total body length measured from anterior margin of vertex to tip of folded fore wing, HW = head width, AL = antennal length, TW = mesoscutum width, WL = fore wing length, and TL = metatibial length. For fifth instar immatures: BL = total body length, HW = head width, AL = antennal length, and FL = fore wing pad length, measured as the distance between transverse tangents of anterior angle and posterior margin. Photos were taken with an Olympus BX41 microscope (Tokyo, Japan) attached to a Cannon 7D camera (Tokyo, Japan). Helicon Focus version 5.3 (Helicon Soft Ltd., Kharkiv, Ukraine) was used for image stacking. The unguitractor of immatures was drawn using Illustrator 2020 (Adobe Systems Inc., San Jose, CA, USA). Line drawings of antenna, male terminalia, paramere, and circumanal ring pore field were undertaken with an Olympus BX41 microscope. Photoshop 2020 (Adobe Systems Inc., USA) was used to edit photos including adjusting background color and cropping without modifying any characters of specimens. All specimens are preserved in the Entomological Museum of China Agricultural University (CAU), Beijing, China. The terminology mainly follows those of Vondráček [20], Ouvrard et al. [21], and Yang et al. [22] for adults and that of White and Hodkinson [23] for immatures. The plant names follow POWO [24].
2.2. Sampling and DNA Extraction
Voucher specimens of Cacopsylla fuscicella sp. nov. were collected on loquat, Eriobotrya japonica, in Zhaotong City, Yunnan Province, China. Most specimens were stored in a refrigerator at −20 °C in 100% ethanol. After being identified based on morphological characters, their genomic DNA was extracted using Dneasy Blood and Tissue Kit (Qiagen, Beijing, China) following the manufacturer’s protocol. Some adult specimens were dry mounted. All specimens are preserved in the Entomological Museum of China Agricultural University (CAU), Beijing, China.
2.3. Mitogenome Sequencing, Assembly, and Annotation
The COX1 fragment (~610 bp) was amplified by polymerase chain reaction (PCR) with newly designed primers UCOIF (5′-TTTCHACNAACCATAAGGAYATTGG-3′) and UCOIR (5′-TANACTTCTGGGTGTCCAAAAAATCA-3′). Short PCR amplifications were carried out using Qiagen Taq DNA polymerase (Qiagen, Beijing, China) with the following cycling conditions: 5 min at 94 °C, followed by 35 cycles of 50 s at 94 °C, 50 sec at 48–55 °C, and 1–2 min at 72 °C. The final elongation step was continued for 10 min at 72 °C. The PCR products were analyzed by 1.0% agarose gel electrophoresis and then sequenced by Sanger sequencing at Sangon Biotech (Beijing, China).
Libraries setting and sequencing were made by the Beijing Genomics institution (Beijing, China). The BGISEQ-500 library was prepared using genomic DNA with an average insert size of 200–400 bp, and pair-end sequencing with 150 bp in length was performed using the BGISEQ-500 sequencer with the processed libraries. For each library, 3 Gb of clean data were obtained after removing reads containing adaptor contamination poly-Ns (>15 bp Ns) or >75 bp bases with quality score ≤ 3. Clean reads were used in de novo assembly by using IDBA-UD [25], with minimum and maximum k-values of 41 and 141 bp, respectively. To identify the corresponding mitogenome assemblies, the assembled contigs were searched with COX1 sequence using BLAST with at least 98% similarity. To investigate the accuracy of the assembly, clear reads were mapped onto the obtained mitogenome sequences using Geneious prime 2022.2.2 (http://www.geneious.com/ accessed on 24 April 2022). Offering four species of Cacopsylla as reference, the mitochondrial genome was extracted and annotated using MitoZ [26]. The annotation accuracy of 13 protein coding genes and 2 rRNA genes were examined by Geneious prime. The online tRNAscan-SE service (http://lowelab.ucsc.edu/tRNAscan-SE/ accessed on 24 April 2022) was used to confirm the locations of the tRNA genes. The complete mitogenome has been uploaded to GenBank (Table 1).
2.4. Phylogenetic Analysis
Combining the newly sequenced mitogenome and sequences from GenBank, a total of 12 species from the family Psyllidae were included in our phylogenetic analysis. Two species, Trioza urticae from Triozidae and Calophya schini from Calophyidae, were selected as outgroups (Table 1).
The two rRNA genes of each species were separately aligned using the L-INS-I strategy in the MAFFT algorithm [27] and trimmed using trimAl v1.2 [28]. The 13 PCGs of each species were translated into amino acids and then aligned using the MAFFT algorithm individually. The sequences of amino acids were trimmed and back translated based on the original DNA sequences in trimAl v1.2. Thirteen PCGs and two rRNA genes were concatenated using FASconCAT-G v 1.04 [29], while the partition file was obtained. Finally, the dataset of PCGRNA matrix with 12,597 nucleotides was conducted.
Phylogenetic trees constructed using the Maximum Likelihood (ML) method were implemented in the IQ-TREE web server with a dataset partitioned by genes [30]. The robustness of the tree topology was further assessed by SH-aLRT, and 1000 ultrafast bootstrap replicates were implemented. The models were determined as follows: TIM+F+I+G4 (for ATP6 and COX2), HKY+F+G4 (for ATP8), TIM2+F+I+G4 (for COX1 and COX3), TPM3u+F+I+G4 (for srRNA), TVM+F+I+G4 (for ND2, ND4, ND4L, ND5 and lrRNA), GTR+F+I+G4 (for CYTB and ND1), HKY+F+I+G4 (for ND3), and TPM3+F+G4 (for ND6).
2.5. Genetic Distance Analysis
We downloaded COX1 sequences of 30 Cacopsylla species, 4 Psylla species, 2 Arytainilla species, and 1 Cyamophila species from GenBank together with the COX1 sequence of the new species, Cacopsylla fuscicella sp. nov., to calculate the genetic distances within Psyllinae (Table S2). The analysis was conducted in MEGA7 [31]. Sequences alignment was performed and positions containing gaps and missing data were eliminated. There was a total of 435 positions in the final dataset. Pairwise distances were calculated based on the p-distance model.
3. Results
3.1. Taxonomy
Suborder: Sternorrhyncha
Superfamily: Psylloidea
Family: Psyllidae Latreille, 1807
Genus: Cacopsylla Ossiannilsson, 1970
Cacopsylla fuscicella sp. nov.
3.1.1. Diagnosis
Cacopsylla fuscicella sp. nov. can be identified by the following combination of characters: genal processes short, about half of median suture of vertex in length; moderately divergent apically, with apices nearly truncate; a width of head about twice as long as the length, the inner-anterior angle of the genae convex.
3.1.2. Description
Adult. Coloration. Summer form (Figure 1A,B): Body orange in the overall view. Vertex mostly orange with white anterior angles. Discal foveae light brown in color. Genal process yellow and lighter than body color. Compound eyes yellow. Lateral ocelli and medial ocellus orange. Occiput black, the postocular sclerite white with black margins. Antennae yellow, with segment ten entirely black. Thoracic dorsum orange in ground with white narrow stripes; pronotum four white markings; one longitudinal white stripe present through the middle of the mesopraescutum. Legs yellow, irregular markings on femora darker. Fore wing membrane hyaline, apices of cells m1, m2 and cu1 with ocherous clouding. Veins near yellow, slightly darker than membrane. Hind wing membrane translucent, more or less brownish along veins; veins light brown to brown. Abdominal terga dark brown with margins of segments III–VI white and segment VI yellowish. Male terminalia orange. Female proctiger ochreous, subgenital plate white with irregular dark brown spots.
Winter form (Figure 1C,D): overall color ochreous, darker than summer form. Fore wing with apical halves of cells r1, r2, m1, m2, cu1, and cu2, dark brown. Posterior margin of each abdominal tergite dark brown.
Structures. Head inclined from the longitudinal body axis by 80–90°, slightly wider than mesoscutum (Figure 1). Lateral parts of vertex short longitudinally; antero-inner angles of vertex produced and rounded; areas around lateral ocelli strongly convex, appearing distinctly contrasted from areas around discal foveae; vertex and gena clearly separated by a suture (Figure 2A). Discal foveae relatively shallow (Figure 2A). Surface of the vertex finely sculptured with scaly microstructuring and microscopic setae (Figure 2A). Genal processes short, at about half of the median suture of vertex in length; moderately divergent apically, with apices being nearly truncated (Figure 2A). Antennae relatively slender, longer than head width, with a single subapical rhinarium on each of segments 4, 6, 8, and 9; the distally situated terminal seta only slightly shorter than the proximally situated terminal seta (Figure 2B). Mesopraescutum moderately protruding forward to force pronotum to arch (Figure 1B,D). Genual spine of metatibia relatively small, apical spurs of metatibia arranged in 1+1+2+1 (Figure 1B). Fore wing oblong oval, widest in apical 1/3; surface spinules covering most of the fore wing membrane, leaving narrow spinule-free bands along veins (Figure 2C).
Male terminalia. Whole proctiger gently arched in lateral aspect, covered with near-evenly spaced setae (Figure 2D). Paramere stout, gently curved; apex of paramere axe-like, curved inwards, with two lateral tips better sclerotized (Figure 2D,E). Inner surface of paramere with many erect short setae (Figure 2E). Distal segment of aedeagus relatively long, slender, and nearly straight; apical dilatation small, near oval, and not hooked; sclerotized end tube of ductus ejaculatorius projected backwards and gently curved upwards (Figure 2D). Subgenital plate subtriangular in profile (Figure 2D).
Female terminalia. The shape is relatively long and simple. The anal ring has a length of about 2/5 of the total length of the proctiger; the dorsum of the proctiger is slightly sinuous in profile. The subgenital plate is relatively long and narrow in profile. Valvulae dorsails and ventralis gently curve upwards and lack teeth (Figure 2F).
Fifth instar immature. Coloration. Generally yellow to orange in color. Membranous parts orange. All sclerites dark brown. Compound eyes red.
Structures. Dorsum completely covered with minute to long simple setae, including ocular and postocular setae (Figure 3A,B). All bases of setae strongly sclerotized. Antenna 8-segmented, with one rhinarium on each apex of segments 4 and 6, and two rhinaria on segment 8 (Figure 3B). Fore wing pad oval in chape, with about ten long setae on the outer margin, a knife-shaped hind wing pad with two long setae on the apex (Figure 3A,B). Abdominal ventrum with long or short conical setae near the margin; other parts with long or short simple setae (Figure 3C). Abdominal margin with rather long 7+7 setae and 3+3 sectasetae (Figure 3C). Circum anal pore field ventral, composed of a complete outer ring of a single row of slit-shaped pores and a complete inner ring of a jagged single row of oval pores (Figure 3D,E). Dorsum of the fore tibial-tarsus with one long seta; dorsum of mid and hind tibial-tarsus with two long setae (Figure 3A,B). Tarsal arolium broadly fishtail shaped with expanded areas rather coarse; pedicel relatively short, gradually narrows basally, and with a well-developed unguitractor (Figure 3F).
3.1.3. Measurements
Measurements are in mm. Adult: ♂ (n = 5): BL 3.24–3.51, AL 1.20–1.26, HW 0.89–0.93, TW 0.91–0.96, WL 2.63–2.82, TL 0.62–0.69. ♀ (n = 4): BL 3.66–3.87, AL 1.22–1.26, HW 0.87–0.90, TW 0.86–0.89, WL 3.06–3.24, TL 0.64–0.70. Immature: (n = 3). BL 1.94–2.18, HW 0.79–0.82, AL 0.83–0.86, FL 0.79–0.85.
3.1.4. Material Examined
Holotype: male, dry mounted, Zhaotong City, Yunnan Province, China, 30 October 2022, Shi Anxian (CAU). Paratypes: 2 males, 2 females, 1 immature, dry mounted, same data as the holotype; 2 males, 2 females, 1 immature, dry mounted, Zhaotong City, Yunnan Province, China, 13 May 2022 (CAU).
3.1.5. Distribution
China: Yunnan Province. Known only from the type locality for now.
3.1.6. Host Plant and Damage
The host plant of this new species Eriobotrya japonica, which is commonly known as loquat. This plant is native to China and is grown commercially for its fruit. As one of the insect pests in the loquat industry in China, it caused great economic losses to fruit growers. They always aggregate on the undersurface of leaves, stem, and bud, sucking phloem sap, wilting the young leaves and flowers, and causing fruit russet (Figure 4).
3.1.7. Etymology
This species is named after its ocherous cells on the forewings of the summer form.
3.1.8. Comments
Before the present study, there have been three psyllid species recorded feeding on the plant genus Eriobotrya [18,19]. Two among them, Cacopsylla eriobotryacola and C. eriobotryae, are the pests of Eriobotrya deflexa. They have yellowish forewings without ocherous clouding and long and thin parameres. Their genae processes are long and have the same length as the median suture of the vertex. For C. eriobotryacola, the gena is thin and acute apically, which can be clearly distinguished from C. fuscicella sp. nov. The other species, C. biwa, is more similar to C. fuscicella sp. nov., both in morphological characters and host plant. They both feed on loquat and Eriobotrya japonica and have a similar color pattern; however, there are still some differences. The head of C. fuscicella sp. nov. is wider than that of C. biwa. The width of the head is about twice as long as the length of head in C. fuscicella sp. nov., but the length–width ratio of the head for C. biwa is lower than two. For C. fuscicella sp. nov., the inner-anterior angle of the genae is convex, but that of C. biwa is flat. The genae of C. fuscicella sp. nov. are shorter and nearly truncate apically. The setae on the caudal plate of C. fuscicella sp. nov. are conspicuously longer than those of C. biwa. In the summer form, the color on the cell of r2 of C. fuscicella is lighter than that of C. biwa. A key to Cacopsylla species associated with Eriobotrya (Rosaceae) was offered as follows.
Key to adults of Cacopsylla species associated with Eriobotrya (Rosaceae)
| 1 | Genae thin and acute apically | C. eriobotryacola |
| - | Not as above | 2 |
| 2 | Genae square, male proctiger only weakly curved proximally | C. eriobotryae |
| - | Genae rounded, male proctiger strongly curved proximally | 3 |
| 3 | The length-width ratio of the head less than 2, the inner-anterior angle of the genae flat | C. biwa |
| - | Width of head about twice as long as the length, the inner-anterior angle of the genae convex | C. fuscicella sp. nov |
3.2. General Features of Cacopsylla fuscicella sp. nov. Mitogenome
The mitogenome of Cacopsylla fuscicella sp. nov. was obtained successfully with a length of 14,832 bp, including 37 coding genes (13 PCGs, 2 rRNA genes, and 22 tRNA genes) and a putative control region (CR) (Table 2, Figure S1). Twenty-three genes were transcribed from the majority strand (J-strand) and the other fourteen genes were transcribed from the minority strand (N-strand). The gene order and orientation were identical with the hypothetical ancestral mitogenome without gene rearrangement. Eight gene overlaps comprising a total of thirty-three bp were identified. In addition to the CR, we also observed 10 non-coding regions ranging from 1 to 29 bp (Table 2). The entire mitogenome was biased towards A and T, with the AT content being 70.9%. The nucleotide skew analysis revealed a positive AT skew (0.075) and a negative CG skew (−0.306) (Table S1).
The total length of the 13 PCGs is 10,768 bp. The protein coding genes have 4 types of start codons (6 ATGs, 4 ATAs, 2 TTGs and 1 ATC), and most started with ATN, except ND5 and ND4L. Nine PCGs stopped with TAA or TAG, while COX2, COX3, ND4, and ND5 truncated termination codons with a single T (Table 2). All 22 tRNA genes were detected and found to be discontinuously scattered throughout the mitogenome. The size of each tRNA gene ranges from 54 bp to 70 bp. The lrRNA gene was 1,102 bp long and was found between trnL1 and trnV, and the srRNA gene, 774 bp in size, was located between trnV and the control region (Table 2, Figure S1). The control region of Cacopsylla fuscicella sp. nov. located between srRNA and trnI was 711 bp in length with the highest AT content (79.1%) in the entire mitogenome (Figure S1). There were no notable repeat elements found in the control region.
3.3. Phylogenetic Analyses and Genetic Distance Analyses
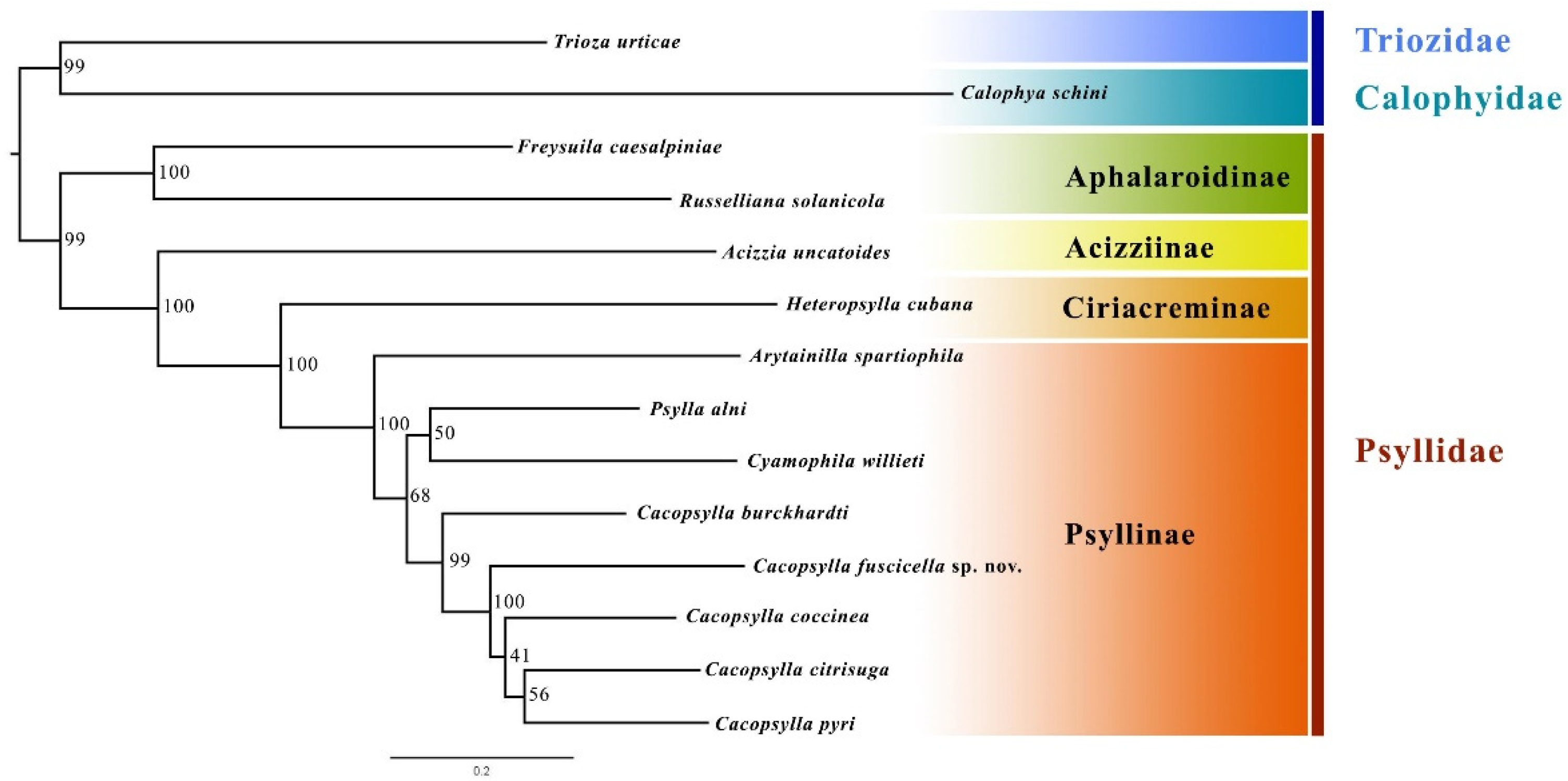
The phylogenetic topology of Psyllidae inferred by the ML method is shown in Figure 5. The monophyly of Aphalaroidinae and Psyllinae was recovered with high support values, and Aphalaroidinae was recovered as the sister group to the other three subfamilies. The results further confirm that the new species described in the present study should be placed in the genus Cacopsylla. The new species, Cacopsylla fuscicella sp. nov., was grouped together with the other four Cacopsylla species, and the sister relationship between C. fuscicella sp. nov. and the clade (C. coccinea + (C. citrisuga + C. pyri)) was strongly supported (node support value = 100).
The results of the genetic distance analysis (Table S2) showed that the average interspecific genetic distance was 0.171 among the 31 Cacopsylla species, 0.161 among the four Psylla species, and 0.177 between the 2 Arytainilla species. The intergeneric distance within Psyllinae was relatively higher with an average value of 0.194 (Arytainilla-Cacopsylla: 0.193, Arytainilla-Cyamophila: 0.189; Arytainilla-Psylla: 0.219; Cacopsylla-Cyamophila: 0.192; Cacopsylla-Psylla: 0.191; Cyamophila-Psylla: 0.178). As for the new species, the average genetic distance between Cacopsylla fuscicella sp. nov. and the other 30 Cacopsylla species was 0.204. However, it is 0.224, 0.221, and 0.216 between Cacopsylla fuscicella sp. nov. and Psylla, Arytainilla, and Cyamophila, respectively.
4. Discussion
Based on morphological characters, Cacopsylla fuscicella sp. nov. can be differentiated from other loquat psyllids by its rounded gena, convex inner anterior angle of genae, and wide head. The degree of DNA barcode also supports the classification of Cacopsylla fuscicella sp. nov. as a new species. It is interesting that the divergence between Cacopsylla fuscicella sp. nov. and C. eriobotryae is not less than divergence between Cacopsylla fuscicella sp. nov. and other Rosaceae-feeding species, which suggests that the two species on Eribotrya are not particularly closely related. Moreover, the divergence between Cacopsylla fuscicella sp. nov. and C. eriobotyrae is greater than that between certain two species from different genera, such as Arytainilla gredi (Ramirez Gomez, 1956) and Psylla buxi (Linnaeus, 1758). This suggests that COI divergence is likely becoming saturated and not a reliable tool for phylogenetic analysis. Therefore, the complete mitochondrial genome is relatively more reliable for confirming systematic placement. Based on the phylogenetic tree we constructed, Cacopsylla fuscicella sp. nov. was grouped within the monophyletic Cacopsylla clade with high support. The relationship of these four subfamilies is congruent with previous phylogenetic analyses of Psylloidea [5].
Psyllids pose a significant threat to crop production and the health of fruit trees, resulting in considerable economic losses. They can deplete plant nutrients, cause tissue damage, and transmit plant pathogens—all of which can have a severe impact on agriculture. The new species, feeding on Eriobotrya japonica, was wildly discovered in the loquat orchard in its type locality. This species can cause damage to foliage and flowers in both the immature and adult stages, as well as impede fruit development and cause injury. Therefore, the production and quality of loquat suffered a strong impact, and it causes damage to the industry of loquat. However, there have been insufficient studies conducted on this species, particularly in the area of taxonomy, which hinders efforts to control the pests and conduct further research. Thus, our study may fill the void by providing morphological and molecular data of Cacopsylla fuscicella sp. nov. Our work was expected to help investigate the biological characteristics of this species, which in turn would enable us to more accurately monitor the population of the pest. With this information, we could develop more effective control strategies and improve our ability to manage the pest population, ultimately reducing the damage it causes to loquats and other affected areas.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/insects14050414/s1, Figure S1. Mitochondrial genome of Cacopsylla fuscicella sp. nov. sequenced in this study. Circular maps were drawn with Geneious prime 2022.2.2 (http://www.geneious.com/ accessed on 4 April 2022). The transcriptional direction is indicated with arrows. Abbreviations: ATP6 and ATP8 for adenosine triphosphate (ATP) synthase subunits 6 and 8; COX1–COX3 for cytochrome oxidase subunits 1–3; CYTB for cytochrome b; ND1–6 and ND4L for nicotinamide adenine dinucleotide hydrogen (NADH) dehydrogenase subunits 1–6 and 4L; lrRNA and srRNA for large and small rRNA subunits; trnX (where X is replaced by one letter amino acid code of the corresponding amino acid), for transfer RNA. Table S1. The nucleotide composition of the Cacopsylla fuscicella sp. nov. mitochondrial genome; Table S2. Genetic distances calculated using p-distance model for COX1 sequences.
Author Contributions
Conceptualization, F.S.; Methodology, Y.Z. and F.S.; Writing—original draft preparation, Y.Z. and A.S.; Writing—review and editing, X.L., Y.L., Y.Z. and W.C.; Visualization, Y.Z., X.L., Y.L. and A.S.; Supervision, F.S.; Funding acquisition, Y.Z. and F.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (Nos. 32120103006, 32170474), the 2115 Talent Development Program of the China Agricultural University and Expert Workstation in Zhaotong, Yunnan (Nos. 2019ZTYX03, 2021ZTYX05).
Data Availability Statement
The mitochondrial genome of Cacopsylla fuscicella sp. nov. has deposited in the GenBank under the accession number OQ390047.
Acknowledgments
We appreciate Zhixin He (China Agricultural University) for his contribution to the illustrations and review of this manuscript. We sincerely thank Yunfei Wu (China Agricultural University) for his kind help with assembling the mitogenome.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hodkinson, I. The biology of the Psylloidea (Homoptera): A review. Bull. Entomol. Res. 1974, 64, 325–338. [Google Scholar] [CrossRef]
- Ossiannilsson, F. Contributions to the knowledge of Swedish Psyllids (Hemiptera: Psyllidae). Entomol. Scand. 1970, 1, 135–144. [Google Scholar] [CrossRef]
- Psyl’list—The World Psylloidea Database. Available online: http://www.Hemiptera-databases.com/psyllist (accessed on 16 December 2022).
- Percy, D.M.; Crampton-Platt, A.; Sveinsson, S.; Lemmon, A.R.; Lemmon, E.M.; Ouvrard, D.; Burckhardt, D. Resolving the psyllid tree of life: Phylogenomic analyses of the superfamily Psylloidea (Hemiptera). Syst. Entomol. 2018, 43, 762–776. [Google Scholar] [CrossRef]
- Cho, G.; Malenovský, I.; Lee, S. Higher-level molecular phylogeny of jumping plant lice (Hemiptera: Sternorrhyncha: Psylloidea). Syst. Entomol. 2019, 44, 638–651. [Google Scholar] [CrossRef]
- Burckhardt, D.; Hodkinson, I. A revision of the west Palaearctic pear psyllids (Hemiptera: Psyllidae). Bull. Entomol. Res. 1986, 76, 119–132. [Google Scholar] [CrossRef]
- Luo, X.; Li, F.; Ma, Y.; Cai, W. A revision of Chinese pear psyllids (Hemiptera: Psylloidea) associated with Pyrus ussuriensis. Zootaxa 2012, 3489, 58–80. [Google Scholar] [CrossRef]
- Cho, G.; Malenovský, I.; Burckhardt, D.; Inoue, H.; Lee, S. DNA barcoding of pear psyllids (Hemiptera: Psylloidea: Psyllidae), a tale of continued misidentifications. Bull. Entomol. Res. 2020, 110, 521–534. [Google Scholar] [CrossRef]
- Song, X.; He, Y.; Wang, X.; Gu, X. The complete mitochondrial genome of Cyamophila willieti (Wu) (Hemiptera: Psyllidae). Mitochondrial DNA B 2019, 4, 3758–3759. [Google Scholar] [CrossRef]
- Jo, E.; Cho, G. The complete mitochondrial genome of Cacopsylla burckhardti (Hemiptera, Psylloidea, Psyllidae). Biodivers Data J. 2022, 10, e85094. [Google Scholar] [CrossRef]
- Que, S.; Yu, L.; Xin, T.; Zou, Z.; Hu, L.; Xia, B. Complete mitochondrial genome of Cacopsylla coccinae (Hemiptera: Psyllidae). Mitochondrial DNA A 2015, 27, 3169–3170. [Google Scholar] [CrossRef]
- Wang, Y.; Cen, Y.; He, Y.; Wu, Y.; Huang, S.; Lu, J. The first complete mitochondrial genome sequence of Cacopsylla citrisuga (Yang & Li), a new insect vector of Huanglongbing in Yunnan Province, China. Mitochondrial DNA B 2021, 6, 575–577. [Google Scholar]
- Kang, A.R.; Kim, M.J.; Park, J.S.; Seo, H.-J.; Song, J.-H.; Won, K.-H.; Choi, E.D.; Kim, I. Comparative analysis of two pear pests, Cacopsylla jukyungi and Cacopsylla burckhardti (Hemiptera: Psyllidae), based on complete mitochondrial genomes and comparison to confamilial species. Agronomy 2022, 12, 2037. [Google Scholar] [CrossRef]
- Yang, C.; Li, F. The pear Psylla (Homoptera) of China with descriptions of seven new species. Entomotaxonomia 1981, 3, 35–47. [Google Scholar]
- Föerster, A. Uebersicht der Gattungen und Arten in der Familie der Psylloden. Verh Nat. Ver Preuss. Rheinl 1848, 5, 65–98. [Google Scholar]
- Hildebrand, M.; Dickler, E.; Geider, K. Occurrence of Erwinia amylovora on insects in a fire blight orchard. J. Phytopathol. 2000, 148, 251–256. [Google Scholar] [CrossRef]
- Cho, G.; Burckhardt, D.; Inoue, H.; Luo, X.; Lee, S. Systematics of the east Palaearctic pear psyllids (Hemiptera: Psylloidea) with particular focus on the Japanese and Korean fauna. Zootaxa 2017, 4362, 75. [Google Scholar] [CrossRef]
- Yang, C.T. Psyllidae of Taiwan. Taiwan Mus. Spec. Publ. Ser. 1984, 3, 1–305. [Google Scholar]
- Inoue, H.; Nakanishi, T.; Kaneda, T. Cacopsylla biwa sp. nov. (Hemiptera: Psyllidae): A new pest of loquat Eriobotrya japonica (Rosaceae) in Japan. Appl. Entomol. Zool. 2013, 49, 11–18. [Google Scholar] [CrossRef]
- Vondráček, K. Mery-Psylloidea (Fauna CSR, Svazek 9); Československá Akademie Věd: Praha, Czech Republic, 1957; p. 451. [Google Scholar]
- Ouvrard, D.; Bourgoin, T.; Campbell, B.C. Comparative morphological assessment of the psyllid pleuron (Insecta, Hemiptera, Sternorrhyncha). J. Morphol. 2002, 252, 276–290. [Google Scholar] [CrossRef]
- Yang, M.; Burckhardt, D.; Fang, S. Psylloidea of Taiwan; National Chung Hsing University: Taichung, China, 2009; Volume 1, p. 96. [Google Scholar]
- White, I.M.; Hodkinson, I.D. Nymphal taxonomy and systematics of the Psylloidea (Homoptera). Bull. Br. Mus. Nat. Hist. (Ent.) 1985, 50, 153–301. [Google Scholar]
- POWO. Plants of the World Online. Available online: http://www.plantsoftheworldonline.org/ (accessed on 4 April 2022).
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA-UD: A de novo assembler for single cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Meng, G.L.; Li, Y.Y.; Yang, C.T.; Liu, S.L. MitoZ: A toolkit for animal mitochondrial genome assembly, annotation and visualization. Nucleic. Acids. Res. 2019, 47, e63. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple sequence alignment software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Kück, P.; Longo, G.C. FASconCAT-G: Extensive functions for multiple sequence alignment preparations concerning phylogenetic studies. Front. Zool. 2014, 11, 81. [Google Scholar] [CrossRef] [PubMed]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast-online phylogenetic tool for maximum likelihood analysis. Nucleic. Acids. Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Dry-mounted specimens of Cacopsylla fuscicella sp. nov. (A). Lateral view of summer form (male). (B). Dorsal view of summer form (male) (C). Lateral view of winter form (female) (D). Dorsal view of winter form (female). Scale bar = 1 mm.
Figure 1.
Dry-mounted specimens of Cacopsylla fuscicella sp. nov. (A). Lateral view of summer form (male). (B). Dorsal view of summer form (male) (C). Lateral view of winter form (female) (D). Dorsal view of winter form (female). Scale bar = 1 mm.

Figure 2.
(A) Head, frontal aspect; (B) antenna; (C) forewing of summer form; (D) male terminalia, lateral aspect; (E) paramere, inner surface; (F) female terminalia, lateral aspect. Scale bars = 0.2 mm for (A,D,E); 0.4 mm for (B,F); 0.5 mm for (C,E).
Figure 2.
(A) Head, frontal aspect; (B) antenna; (C) forewing of summer form; (D) male terminalia, lateral aspect; (E) paramere, inner surface; (F) female terminalia, lateral aspect. Scale bars = 0.2 mm for (A,D,E); 0.4 mm for (B,F); 0.5 mm for (C,E).

Figure 3.
(A) fifth instar immature, ventral aspect; (B) fifth instar immature, dorsal aspect; (C) abdominal sectaseta; (D,E) circumanal ring, with an outer and inner ring of pores. F. tarsal arolium; Scale bars = 0.5 mm for (A–C); 0.4 mm for (D); 0.2 mm for (E); 0.025 mm for (F).
Figure 3.
(A) fifth instar immature, ventral aspect; (B) fifth instar immature, dorsal aspect; (C) abdominal sectaseta; (D,E) circumanal ring, with an outer and inner ring of pores. F. tarsal arolium; Scale bars = 0.5 mm for (A–C); 0.4 mm for (D); 0.2 mm for (E); 0.025 mm for (F).

Figure 4.
Damage of Cacopsylla fuscicella sp. nov. to Eriobotrya japonica. (A) Nymphs aggregating on base of young leaves; (B) Nymphs aggregating on stem; (C) Nymphs aggregating on bud; (D) Adults aggregating on the undersurface of leaves; (E) Damage to flowers; (F) Damage to fruits.
Figure 4.
Damage of Cacopsylla fuscicella sp. nov. to Eriobotrya japonica. (A) Nymphs aggregating on base of young leaves; (B) Nymphs aggregating on stem; (C) Nymphs aggregating on bud; (D) Adults aggregating on the undersurface of leaves; (E) Damage to flowers; (F) Damage to fruits.

Figure 5.
Phylogenetic relationship of Psyllidae inferred via IQ-TREE based on mitochondrial genome sequences. Numbers close to the branching points are bootstrap support values.
Figure 5.
Phylogenetic relationship of Psyllidae inferred via IQ-TREE based on mitochondrial genome sequences. Numbers close to the branching points are bootstrap support values.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Taxa used in phylogenetic analysis.
| Family | Subfamily | Species | Genbank Accession Number |
|---|---|---|---|
| Triozidae | Triozinae | Trioza urticae (Linné, 1758) | NC_038113 |
| Calophyidae | Calophyinae | Calophya schini Tuthill, 1959 | MF431591 |
| Psyllidae | Aphalaroidinae | Freysuila caesalpiniae (Tuthill, 1959) | MG989225 |
| Russelliana solanicola (Tuthill, 1959) | NC_038140 | ||
| Acizziinae | Acizzia uncatoides (Ferris & Klyver, 1932) | NC_038146 | |
| Ciriacreminae | Heteropsylla cubana (Crawford, 1914) | MG989226 | |
| Psyllinae | Arytainilla spartiophila (Foerster, 1848) | NC_038133 | |
| Psylla alni (Linné, 1758) | NC_038139 | ||
| Cyamophila willieti (Wu, 1932) | MN364946 | ||
| Cacopsylla burckhardti (Luo, Li, Ma & Cai, 2012) | OK574466 | ||
| Cacopsylla fuscicella sp. nov. | OQ390047 | ||
| Cacopsylla coccinea (Kuwayama, 1908) | NC_027087 | ||
| Cacopsylla citrisuga (Yang & Li, 1984) | NC_053749 | ||
| Cacopsylla pyri (Linné, 1758) | NC_038148 |
Table 2.
Organization of Cacopsylla fuscicella sp. nov. mitochondrial genome.
| Gene | Direction | Location | Size (bp) | Anticodon | Codon Start | Codon Stop | Intergenic Nucleotides |
|---|---|---|---|---|---|---|---|
| tRNAIle | F | 1–66 | 66 | GAT | |||
| tRNAGln | R | 71–136 | 66 | TTG | 4 | ||
| tRNAMet | F | 143–208 | 66 | CAT | 6 | ||
| ND2 | F | 209–1180 | 972 | ATG | TAA | 0 | |
| tRNATrp | F | 1203–1267 | 65 | TCA | 22 | ||
| tRNACys | R | 1271–1333 | 63 | GCA | 3 | ||
| tRNATyr | R | 1334–1395 | 62 | GTA | 0 | ||
| COI | F | 1397–2929 | 1533 | ATG | TAA | 1 | |
| tRNALeu(UUR) | F | 2929–2996 | 68 | TAA | −1 | ||
| COII | F | 2997–3660 | 664 | ATA | T-- | 0 | |
| tRNALys | F | 3661–3730 | 70 | CTT | 0 | ||
| tRNAAsp | F | 3737–3802 | 66 | GTC | 6 | ||
| ATP8 | F | 3803–3955 | 153 | ATA | TAG | 0 | |
| ATP6 | F | 3949–4623 | 675 | ATG | TAA | −7 | |
| COIII | F | 4623–5400 | 778 | ATG | T-- | −1 | |
| tRNAGly | F | 5401–5458 | 58 | TCC | 0 | ||
| ND3 | F | 5459–5809 | 351 | ATA | TAG | 0 | |
| tRNAAla | F | 5808–5868 | 61 | TGC | −2 | ||
| tRNAArg | F | 5870–5930 | 61 | TCG | 1 | ||
| tRNAAsn | F | 5931–5997 | 67 | GTT | 0 | ||
| tRNASer(AGN) | F | 5998–6051 | 54 | GCT | 0 | ||
| tRNAGlu | F | 6052–6113 | 62 | TTC | 0 | ||
| tRNAPhe | R | 6102–6164 | 63 | GAA | −12 | ||
| ND5 | R | 6165–7782 | 1618 | TTG | T-- | 0 | |
| tRNAHis | R | 7783–7843 | 61 | GTG | 0 | ||
| ND4 | R | 7844–9086 | 1243 | ATG | T-- | 0 | |
| ND4L | R | 9080–9367 | 288 | TTG | TAG | −7 | |
| tRNAThr | F | 9369–9429 | 61 | TGT | 1 | ||
| tRNAPro | R | 9430–9491 | 62 | TGG | 0 | ||
| ND6 | F | 9494–9979 | 486 | ATC | TAA | 2 | |
| CytB | F | 9979–11,115 | 1137 | ATG | TAG | −1 | |
| tRNASer(UCN) | F | 11,114–11,175 | 62 | TGA | −2 | ||
| ND1 | R | 11,205–12,119 | 915 | ATA | TAA | 29 | |
| tRNALeu(CUN) | R | 12,120–12,183 | 64 | TAG | 0 | ||
| lrRNA | R | 12,184–13,285 | 1102 | 0 | |||
| tRNAVal | R | 13,286–13,347 | 62 | TAC | 0 | ||
| srRNA | R | 13,348–14,121 | 774 | 0 | |||
| Control region | 14,122–14,832 | 711 | 0 |
* Negative numbers indicate that adjacent genes overlap.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhao, Y.; Luo, X.; Liu, Y.; Shi, A.; Cai, W.; Song, F. Cacopsylla fuscicella Sp. Nov. (Hemiptera, Psyllidae), a New Loquat Pest in China. Insects 2023, 14, 414. https://doi.org/10.3390/insects14050414
AMA Style
Zhao Y, Luo X, Liu Y, Shi A, Cai W, Song F. Cacopsylla fuscicella Sp. Nov. (Hemiptera, Psyllidae), a New Loquat Pest in China. Insects. 2023; 14(5):414. https://doi.org/10.3390/insects14050414
Chicago/Turabian StyleZhao, Yisheng, Xinyu Luo, Yingqi Liu, Anxian Shi, Wanzhi Cai, and Fan Song. 2023. "Cacopsylla fuscicella Sp. Nov. (Hemiptera, Psyllidae), a New Loquat Pest in China" Insects 14, no. 5: 414. https://doi.org/10.3390/insects14050414
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.