
Vegetation Types Attributed to Deforestation and Secondary Succession Drive the Elevational Changes in Diversity and Distribution of Terrestrial Mosses in a Tropical Mountain Forest in Southern China

Abstract
:
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Moss Sampling
2.3. Environmental and Climatic Factors
2.4. Statistical Analysis
3. Results
3.1. Species Composition
3.2. Distribution of Terrestrial Mosses along the Elevation Gradient
3.3. Life Form Composition
3.4. Phytogeographical Elements
3.5. Environmental Variables and Species Composition
4. Discussion
5. Conclusions
6. Future Research
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation | Species | Family | Life Form | Geographical Element |
|---|---|---|---|---|
| 100 | Syrrhopodon gardneri | Calymperaceae | Turf | Pantropical |
| Syrrhopodon armatus | Calymperaceae | Turf | Palaeotropical | |
| Hyophlia javanica | Pottiaceae | Turf | Tropical Asian | |
| Philonotis hastata | Bartramiaceae | Cushion | Pantropical | |
| Octoblepharum albidum | Leucobryaceae | Cushion | Pantropical | |
| Schwetschkeopsis fabronia | Fabroniaceae | Weft | North temperate | |
| Claopodium gracillimum | Thuidiaceae | Mat | East Asian | |
| Claopodium pellucinerve | Thuidiaceae | Mat | North temperate | |
| Entodontopsis anceps | Stereophyllaceae | Mat | Tropical Asian | |
| Fissidens guangdongenis | Fissidentaceae | Turf | Tropical Asian | |
| Sematophyllum subpinnatum | Sematophyllaceae | Mat | Pantropical | |
| Pseudotaxiphyllum densum | Hypnaceae | Mat | East Asian | |
| Ectropothecium zollingeri | Hypnaceae | Mat | East Asian-Tropical Asian-Australian-Oceanian | |
| Vesicularia montagnei | Hypnaceae | Mat | East Asian-Tropical Asian-Australian-Oceanian | |
| Pogonatum inflexum | Polytrichaceae | Turf | East Asian Tropical Asian | |
| 300 | Pleuridium subulatum | Ditrichaceae | Turf | North temperate |
| Campylopus laxitextus | Dicranaceae | Cushion | East Asian-Tropical Asian-Australian-Oceanian | |
| Campulopus subulatus | Dicranaceae | Cushion | North temperate | |
| Fissidens guangdongenis | Fissidentaceae | Turf | Tropical Asian | |
| Fissidens crispulus | Fissidentaceae | Turf | Palaeotropical | |
| Leucobryum humillimum | Leucobryaceae | Weft | East Asian Tropical Asian | |
| Duthiella speciosissima | Trachypodaceae | Weft | East Asian | |
| Taxithelium oblongifolium | Sematophyllaceae | Mat | East Asian Tropical Asian | |
| Sematophyllum phoenicium | Sematophyllaceae | Mat | Palaeotropical | |
| Pseudotaxiphyllum pohliaecarpum | Hypnaceae | Mat | East Asian-Tropical Asian-Australian-Oceanian | |
| 500 | Brothera leana | Dicranaceae | Turf | North temperate |
| Campylopus gracilis | Dicranaceae | Cushion | North temperate | |
| Campylopus umbellatus | Dicranaceae | Cushion | East Asian-Tropical Asian-Australian-Oceanian | |
| Dicranodontium didymodon | Dicranaceae | Cushion | Endemic to China | |
| Leucobryum bowringii | Leucobryaceae | Weft | Pantropical | |
| Leucobryum boninense | Leucobryaceae | Weft | East Asian | |
| Octoblepharum albidum | Leucobryaceae | Cushion | Pantropical | |
| Fissidens guangdongenis | Fissidentaceae | Turf | Tropical Asian | |
| Sematophyllum subhumile | Sematophyllaceae | Mat | East Asian-Tropical Asian-Australian-Oceanian | |
| Sematophyllum subpinnatum | Sematophyllaceae | Mat | Pantropical | |
| Papillidiopsis macrosticta | Sematophyllaceae | Cushion | East Asian | |
| Pseudotaxiphyllum pohliaecarpum | Hypnaceae | Mat | East Asian-Tropical Asian-Australian-Oceanian | |
| 700 | Campulopus ericoides | Dicranaceae | Cushion | Tropical Asian |
| Leucobryum scabrum | Leucobryaceae | Weft | East Asian | |
| Leucobryum juniperoideum | Leucobryaceae | Weft | North temperate | |
| Hyophlia involuta | Pottiaceae | Turf | Pantropical | |
| Pohlia nutans | Bryaceae | Turf | Cosmopolitan | |
| Thuidium pristocalyx | Thuidiaceae | Mat | Palaeotropical | |
| Claopodium assurgens | Thuidiaceae | Mat | East Asian-Tropical Asian-Australian-Oceanian | |
| Hypnum plumaeformae var. minus | Hypnaceae | Weft | East Asian | |
| Pseudotaxiphyllum densum | Hypnaceae | Mat | East Asian | |
| Pogonatum neesii | Polytrichaceae | Turf | East Asian-Tropical Asian-Australian-Oceanian | |
| 900 | Campylopus umbellatus | Dicranaceae | Cushion | East Asian-Tropical Asian-Australian-Oceanian |
| Fissidens crispulus | Fissidentaceae | Turf | Palaeotropical | |
| Hyophlia javanica | Pottiaceae | Turf | Tropical Asian | |
| Pseudosymblepharis angustata | Pottiaceae | Cushion | Palaeotropical | |
| Archidium ohioense | Archidiaceae | Turf | Pantropical | |
| Anomobryum julaceum | Bryaceae | Cushion | Cosmopolitan | |
| Plagiomnium vesicatum | Mniaceae | Mat | Pantropical | |
| Pterobryopsis crassicaulis | Pterobryaceae | Mat | Tropical Asian | |
| Chrysocladium retrorsum | Meteoriaceae | Weft | East Asian Tropical Asian | |
| Thuidium glaucinoides | Thuidiaceae | Mat | Tropical Asian | |
| Brachythecium garovaglioides | Brachytheciaceae | Weft | East Asian | |
| Brachythecium buchananii | Brachytheciaceae | Weft | East Asian Tropical Asian | |
| Sematophyllum subpinnatum | Sematophyllaceae | Mat | Pantropical | |
| Hypnum plumaeforme | Hypnaceae | Weft | East Asian Tropical Asian | |
| Hypnum callichroum | Hypnaceae | Weft | North temperate |
Appendix B
| Site No. | Transect No. | Altitude (m asl) | Total Number of Species | Cover (%) | Thickness (mm) |
|---|---|---|---|---|---|
| 1 | 1 | 100 | 10 | 17.0 | 3.1 |
| 2 | 110 | 6 | 14.5 | 2.8 | |
| 3 | 120 | 8 | 13.6 | 3.8 | |
| 2 | 1 | 290 | 10 | 10.2 | 3.5 |
| 2 | 306 | 7 | 10.8 | 3.2 | |
| 3 | 315 | 6 | 10.2 | 3.0 | |
| 3 | 1 | 490 | 7 | 15.8 | 3.1 |
| 2 | 505 | 8 | 10.7 | 3.3 | |
| 3 | 512 | 6 | 10.3 | 4.0 | |
| 4 | 1 | 690 | 7 | 7.3 | 4.5 |
| 2 | 705 | 3 | 7.0 | 6.0 | |
| 3 | 720 | 5 | 7.3 | 4.7 | |
| 5 | 1 | 890 | 6 | 23.5 | 8.1 |
| 2 | 900 | 10 | 25.0 | 10.0 | |
| 3 | 912 | 7 | 23.0 | 7.7 |
Appendix C
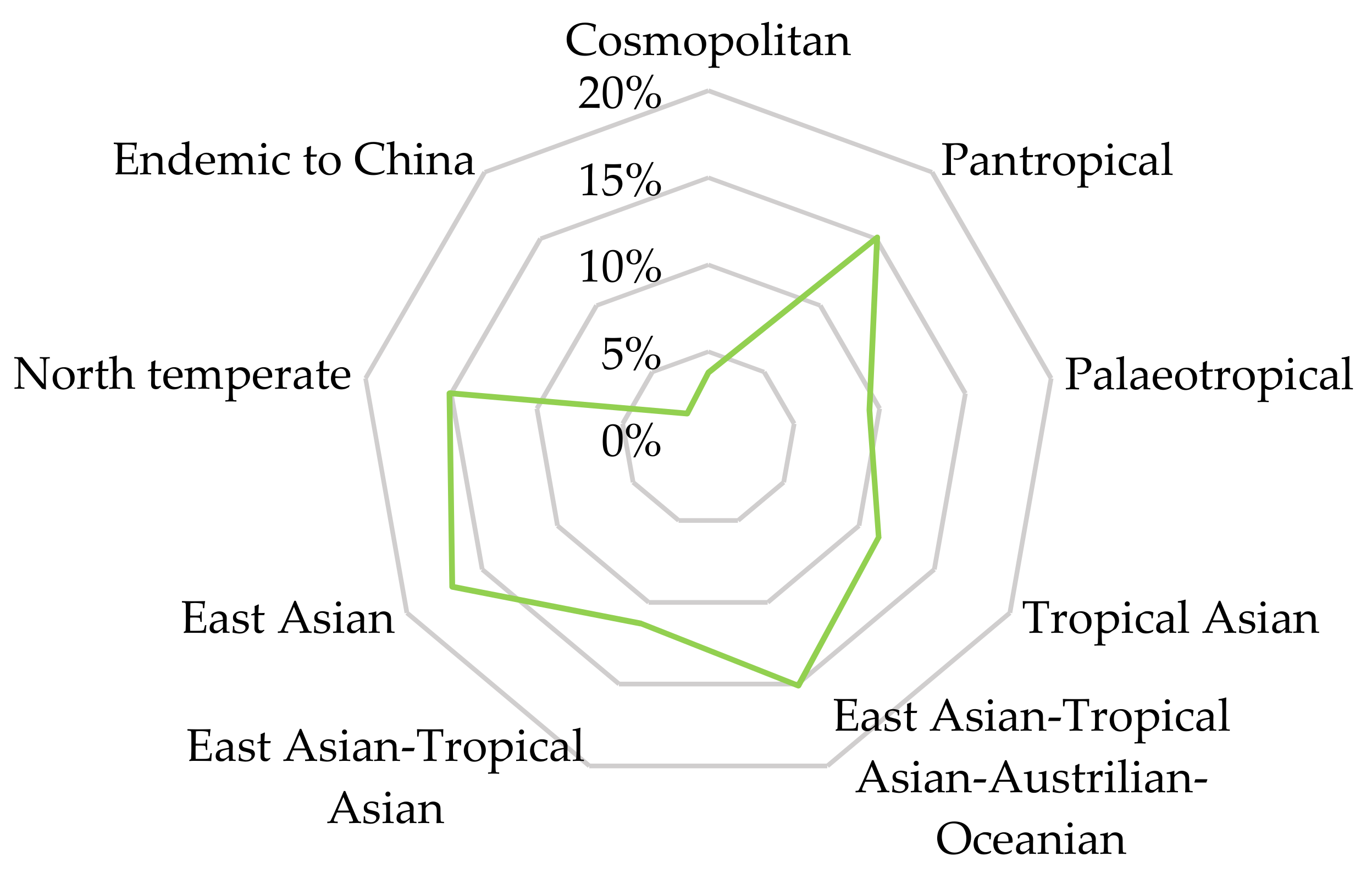
| Patterns | Moss Species (% Total) |
|---|---|
| 1. Cosmopolitan | 2 (3.8%) |
| 2. Pantropical | 8 (15.1%) |
| 3. Paleotropical | 5 (9.4%) |
| 4. Tropical Asian | 6 (11.3%) |
| 5. East Asian–Tropical Asian–Australian–Oceanian | 8 (15.1%) |
| 6. East Asian | 9 (17.0%) |
| 7. East Asian–Tropical Asian | 6 (11.3%) |
| 8. Northern temperate | 8 (15.1%) |
| 9. Endemic to China | 1 (1.9%) |
| Total | 53 |
Appendix D
| Canonical Variable Sets | Canonical Correlation Coefficient | Canonical Variance Explained (%) | F | df | p |
|---|---|---|---|---|---|
| 1 | 0.985 | 84.9 | 8.542 | 15 | 0.000 |
| 2 | 0.922 | 14.6 | 3.533 | 8 | 0.015 |
| 3 | 0.355 | 0.4 | 0.432 | 3 | 0.735 |
Appendix E
| Set 1 | Set 2 | Set 3 | |
|---|---|---|---|
| Biotic variables | |||
| Species richness | 0.267 | 0.617 | 0.741 |
| Cover | 0.961 | 0.274 | −0.030 |
| Thickness | 0.846 | −0.514 | 0.143 |
| Environmental variables | |||
| Air temperature | −0.621 | 0.658 | −0.085 |
| Dew point | −0.800 | 0.486 | −0.088 |
| Relative humidity | 0.352 | −0.736 | 0.051 |
| Rainfall | 0.733 | −0.623 | 0.082 |
| pH | 0.069 | 0.389 | 0.067 |
| Ground cover | −0.232 | 0.878 | −0.035 |
| Canopy closure | −0.453 | 0.845 | −0.086 |
| Wind speed | 0.498 | −0.738 | −0.029 |
| Altitude | 0.566 | −0.750 | 0.048 |
Appendix F
| Canonical Variable Sets | Set 1 by Self | Set 1 by Set 2 | Set 2 by Self | Set 2 by Set 1 |
|---|---|---|---|---|
| 1 | 57.0 | 55.3 | 28.0 | 27.2 |
| 2 | 24.0 | 20.4 | 48.2 | 41.0 |
| 3 | 19.0 | 2.4 | 0.4 | 0.1 |
References
- Goffinet, B.; Buck, W.R. Systematics of Bryophyta: From molecules to a revised classification. In Molecular Systematics of Bryophytes (Monographs in Systematic Botany); Goffinet, B., Hollowel, V., Magill, R., Eds.; Missouri Botanical Garden Press: St. Louis, MO, USA, 2004; pp. 205–239. [Google Scholar]
- Dowle, E.J.; Morgan-Richards, M.; Trewick, S.A. Molecular evolution and the latitudinal biodiversity gradient. Heredity 2013, 110, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Hillebrand, H. On the generality of the latitudinal diversity gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [Green Version]
- Sahney, S.; Benton, M.J. Recovery from the most profound mass extinction of all time. Proc. R. Soc. B Biol. Sci. 2008, 275, 759–765. [Google Scholar] [CrossRef] [Green Version]
- Frahm, J.-P. Diversity of bryophyte species in the tropics. Trop. Bryol. 2003, 23, 13–21. [Google Scholar]
- Gaston, K.J. Global patterns in biodiversity. Nature 2000, 405, 220–227. [Google Scholar] [CrossRef]
- Glime, J.M. Tropics: Disturbance and conservation. In Bryophyte Ecology: Habitat and Role; Glime, J.M., Ed.; Michigan Tech Open Access Publications: Houghton, MI, USA, 2019; Volume 4, pp. 223–232. [Google Scholar]
- Glime, J.M. Meet the bryophytes. In Bryophyte Ecology: Physiological Ecology; Glime, J.M., Ed.; Michigan Tech Open Access Publications: Houghton, MI, USA, 2017; Volume 1, pp. 1–10. [Google Scholar]
- Grau, O.; Grytnes, J.; Birks, H.J.B. A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya. J. Biogeogr. 2007, 34, 1907–1915. [Google Scholar] [CrossRef]
- Grytnes, J.A.; Heegaard, E.; Ihlen, P.G. Species richness of vascular plants, bryophytes and lichens along an altitudinal gradient in western Norway. Acta Oecol. 2006, 29, 241–246. [Google Scholar] [CrossRef]
- Liede-Schumann, S.; Breckle, S.-W. Bryophyta checklists. In Provisional Checklists of Flora and Fauna of the San Francisco Valley and its Surroundings (Reserva Biológica San Francisco, Province Zamora-Chinchipe, Southern Ecuador); Liede-Schumann, S., Breckle, S.-W., Eds.; Society of Tropical Ecology: Bonn, Germany, 2008; pp. 60–89. [Google Scholar]
- Belnap, J.; Budel, B.; Lange, O.L. Biological soil crusts: Characteristics and distribution. In Ecological Studies: Biological Soil Crusts: Structure, Function, and Management; Belnap, J., Lange, O.L., Eds.; Springer: Berlin, Germany, 2003; Volume 150, pp. 3–30. [Google Scholar]
- Seitz, S.; Nebel, M.; Goebes, P.; Käppeler, K.; Schmidt, K.; Shi, X.; Song, Z.; Webber, C.L.; Weber, B.; Scholten, T. Bryophyte-dominated biological soil crusts mitigate soil erosion in an early successional Chinese subtropical forest. Biogeosciences 2017, 14, 5775–5788. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zheng, Y.; Zhang, B.; Chen, Y.; Zhang, Y. Progress in the study of algae and mosses in biological soil crusts. Front. Biol. China 2009, 4, 143–150. [Google Scholar] [CrossRef]
- Clark, K.L.; Nadkarni, N.M.; Gholz, H.L. Retention of inorganic nitrogen by epiphytic bryophytes in a tropical montane forest. Biotropica 2005, 37, 328–336. [Google Scholar] [CrossRef]
- Turetsky, M.R. The role of bryophytes in carbon and nitrogen cycling. Bryologist 2003, 106, 395–409. [Google Scholar] [CrossRef]
- Beringer, J.; Lynch, A.H.; Chapin, F.S.; Mack, M.; Bonan, G.B. The representation of arctic soils in the land surface model: The importance of mosses. J. Clim. 2001, 14, 3324–3335. [Google Scholar] [CrossRef]
- Hölscher, D.; Köhler, L.; van Dijk, A.; Bruijnzeel, L.A. The importance of epiphytes to total rainfall interception by a tropical montane rain forest in Costa Rica. J. Hydrol. 2004, 292, 308–322. [Google Scholar] [CrossRef]
- Delach, A.; Kimmerer, R.W. The effect of Polytrichum piliferum on seed germination and establishment on iron mine tailings in New York. Bryologist 2002, 105, 249–255. [Google Scholar] [CrossRef]
- Jeschke, M.; Kiehl, K. Effects of a dense moss layer on germination and establishment of vascular plants in newly created calcareous grasslands. Flora 2008, 203, 557–566. [Google Scholar] [CrossRef]
- Shen, T.; Corlett, R.T.; Song, L.; Ma, W.-Z.; Guo, X.-L.; Song, Y.; Wu, Y. Vertical gradient in bryophyte diversity and species composition in tropical and subtropical forests in Yunnan, SW China. J. Veg. Sci. 2018, 29, 1075–1087. [Google Scholar] [CrossRef]
- Song, L.; Liu, W.Y.; Nadkarni, N.M. Response of non-vascular epiphytes to simulated climate change in a montane moist evergreen broad-leaved forest in southwest China. Biol. Conserv. 2012, 152, 127–135. [Google Scholar] [CrossRef]
- He, X.; He, K.S.; Hyvönen, J. Will bryophytes survive in a warming world? Perspect. Plant Ecol. Evol. Syst. 2016, 19, 49–60. [Google Scholar] [CrossRef]
- Raabea, S.; Müllerb, J.; Manthey, M.; Dürhammer, O.; Teuber, U.; Göttlein, A. Drivers of bryophyte diversity allow implications for forest management with a focus on climate change. For. Ecol. Manag. 2010, 260, 1956–1964. [Google Scholar] [CrossRef]
- Kricher, J.C. Other tropical ecosystems: From the mountains to the rivers to the sea. In Tropical Ecology; Princeton University Press: Princeton, NJ, USA, 2011; pp. 423–437. [Google Scholar]
- Shields, J.M.; Webster, C.R.; Glime, J.M. Bryophyte community response to silvicultural opening size in a managed northern hardwood forest. For. Ecol. Manag. 2007, 252, 222–229. [Google Scholar] [CrossRef]
- Weibull, H.; Rydin, H. Bryophytes species richness on boulders: Relationship to area, habitat diversity and canopy tree species. Biol. Conserv. 2005, 122, 71–79. [Google Scholar] [CrossRef]
- Bardat, J.; Aubert, M. Impact of forest management on the diversity of corticolous bryophyte assemblages in temperate forests. Biol. Conserv. 2007, 139, 47–66. [Google Scholar] [CrossRef]
- Ah-Peng, C.; Chuah-Petiot, M.; Descamps-Julien, B.; Bardat, J.; Stamenoff, P. Bryophyte diversity and distribution along an altitudinal gradient on a lava flow in La Reunion. Divers. Distrib. 2007, 13, 654–662. [Google Scholar] [CrossRef]
- Andrew, N.R.; Rodgerson, L.; Dunlop, M. Variation in invertebrate-bryophyte community structure at different spatial scales along altitudinal gradients. J. Biogeogr. 2003, 30, 731–746. [Google Scholar] [CrossRef]
- Bruun, H.H.; Moen, J.; Virtanen, R.; Grytnes, J.A.; Oksanen, L. Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. J. Veg. Sci. 2006, 17, 37–46. [Google Scholar] [CrossRef]
- Frahm, J.-P. Ecology of bryophytes along altitudinal and latitudinal gradients in Chile. Trop. Bryol. 2002, 21, 67–79. [Google Scholar]
- Stehn, S.E.; Webster, C.R.; Glime, J.M. Elevational gradients of bryophyte diversity, life forms, and community assemblage in the southern Appalachian Mountains. Can. J. For. Res. 2010, 40, 2164–2174. [Google Scholar] [CrossRef]
- Corlett, R.T. Human impact on the flora of Hong Kong Island. In The Changing Face of East Asia during Tertiary and Quaternary; Jablonski, N., Ed.; Center of Asian Research: Hong Kong, China, 1997; pp. 400–412. [Google Scholar]
- Zhuang, X.Y.; Corlett, R.T. Forest and forest succession in Hong Kong. J. Trop. Ecol. 1997, 14, 857–866. [Google Scholar] [CrossRef] [Green Version]
- Ashworth, J.M.; Corlett, R.T.; Dudgeon, D.; Melville, D.S.; Tang, W.S.M. Hong Kong Ecological Database; World Wide Fund for Nature: Hong Kong, China, 1993. [Google Scholar]
- Corlett, R.T. Environmental heterogeneity and species survival in degraded tropical landscapes. In The Ecological Consequences of Environmental Heterogeneity; Hutchings, M.J., John, E.A., Stewart, A., Eds.; Blackwell Science: Oxford, UK, 2000; pp. 333–355. [Google Scholar]
- Owen, B.; Shaw, R. Hong Kong Landscapes: Shaping the Barren Rock; Hong Kong University Press: Hong Kong, China, 2007. [Google Scholar]
- Planning Department. Land Use in Hong Kong. Available online: https://www.pland.gov.hk/pland_en/info_serv/statistic/landu.html (accessed on 1 September 2018).
- Agriculture, Fisheries and Conservation Department. Hong Kong: The Facts—Country Parks and Special Areas. Available online: https://www.afcd.gov.hk/english/country/cou_lea/the_facts.html (accessed on 11 June 2021).
- Dudgeon, D.; Corlett, R.T. Hills and Streams: An Ecology of Hong Kong; Hong Kong University Press: Hong Kong, China, 1994. [Google Scholar]
- Agriculture, Fisheries and Conservation Department. Hong Kong Biodiversity Database: The Natural Environment, Plants & Animals in Hong Kong. Available online: https://www.afcd.gov.hk/english/conservation/hkbiodiversity/hkbiodiversity.html (accessed on 1 September 2020).
- Hu, Q.-M. Preface: Rare and Precious Plants of Hong Kong; Agriculture, Fisheries and Conservation Department, Hong Kong Government: Hong Kong, China, 2003. [Google Scholar]
- Zhang, L. An updated and annotated inventory of Hong Kong Bryophytes. Mem. Hong Kong Nat. Hist. Soc. 2003, 26, 1–133. [Google Scholar]
- Hong Kong Observatory. Monthly Means of Meteorological Elements for Tai Mo Shan, 1997–2016: Cold/Hot Weather and Rainfall Statistics. Available online: https://www.hko.gov.hk/en/wxinfo/pastwx/mws2016/mws201601.htm (accessed on 2 February 2016).
- BGCI Webinar Series: Increasing Native Species Supply for Ecological Restoration: Restoring a Diverse Forest in Hong Kong. Available online: https://www.bgci.org/news-events/bgci-webinar-series-increasing-native-species-supply-for-ecological-restoration/ (accessed on 27 May 2021).
- Wu, D.L.; Zhang, L. Bryophyte Flora of Guangdong, 1st ed.; Guangdong Science and Technology Press: Guangzhou, China, 2013. [Google Scholar]
- Wu, P.C.; Jia, Y.; Wang, Q.H.; Yu, N.-N.; He, Q.; Wang, M.Z. Chinese Illustrated Bryophytes, 1st ed.; China Forestry Publishing House: Beijing, China, 2018. [Google Scholar]
- Shen, L.; Yu, J.; Li, D.-D.; Guo, S.-L. Geographical elements of bryophytes recorded from Zhoushan archipelago-discussion on methods of geographical element division of bryophytes. Bull. Bot. Res. 2019, 39, 826–834. [Google Scholar]
- Korhonen, L.; Korhonen, K.T.; Rautiainen, M.; Stenberg, P. Estimation of forest canopy cover: A comparison of field measurement techniques. Silva Fenn. 2006, 40, 315. [Google Scholar] [CrossRef] [Green Version]
- Dick, W.A.; Cheng, L.; Wang, P. Soil acid and alkaline phosphatase activity as pH adjustment indicators. Soil Biochem. 2000, 32, 1915–1919. [Google Scholar] [CrossRef]
- Afifi, A.; Clark, V.A.; May, S. Computer-Aided Multivariate Analysis, 4th ed.; Chapman & Hall/CRC Press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Garson, G.D. GLM Multivariate, MANOVA, and Canonical Correlation; Statistical Associates Publishers: Asheboro, NC, USA, 2015. [Google Scholar]
- Wagner, S.; Bader, M.Y.; Zotz, G. Physiological ecology of tropical bryophytes. In Photosynthesis in Bryophytes and Early Land Plants: Advances in Photosynthesis and Respiration; Hanson, D., Rice, S., Eds.; Springer: Dordrecht, The Netherlands, 2014; Volume 37, pp. 269–289. [Google Scholar]
- Zotz, G. Altitudinal changes in diversity and abundance of non-vascular epiphytes in the tropics: An ecophysiological explanation. Selbyana 1999, 20, 256–260. [Google Scholar]
- Wagner, S.; Zotz, G.; Bader, M.Y. The temperature acclimation potential of terrestrial bryophytes. Plant Biol. 2014, 16, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Longton, R.E. Physiological processes and response to stress. In Biology of Polar Bryophytes and Lichens; Longton, R.E., Ed.; Cambridge University Press: Cambridge, UK, 1988; pp. 141–210. [Google Scholar]
- Marchall, M.; Proctor, M.C.F. Are bryophytes shade plants? Photosynthetic light responses and proportions of chlorophyll a, chlorophyll b and total carotenoids. Ann. Bot. 2004, 94, 593–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuba, Z. Light, temperature and desiccation responses of CO2-exchange in desiccation tolerant moss, Tortula ruralis. Symp. Biol. Hung. 1987, 35, 137–150. [Google Scholar]
- Corrales, A.; Duque, A.; Uribe, J.; Londoño, V. Abundance and diversity patterns of terrestrial bryophyte species in secondary and planted montane forests in the northern portion of the Central Cordillera of Colombia. Bryologist 2010, 113, 8–21. [Google Scholar] [CrossRef]
- Peintinger, M.; Bergamini, A. Community structure and diversity of bryophytes and vascular plants in abandoned fen meadows. Plant Ecol. 2006, 185, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Virtanen, R.; Crawley, M.J. Contrasting patterns in bryophyte and vascular plant species richness in relation to elevation, biomass and Soay sheep on St Kilda, Scotland. Plant Ecol. Divers. 2010, 3, 77–85. [Google Scholar] [CrossRef]
- Proctor, M.C.F. Climate responses and limits of bryophytes: Comparisons and contracts with vascular plants. In Bryophyte Ecology and Climate Change; Slack, N., Tuba, Z., Stark, L.R., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 35–54. [Google Scholar]
- Porley, R.; Hodgetts, N. Mosses and Liverworts; Harper Collins Publishers: London, UK, 2005. [Google Scholar]
- Spitale, D. The interaction between elevation gradient and substratum reveals how bryophytes respond to the climate. J. Veg. Sci. 2016, 27, 844–853. [Google Scholar] [CrossRef]
- Wang, Z.-M.; Ye, W.; Xing, F.-W. Bryophyte diversity on a tropical continental island (Hainan, China): Potential vulnerable species and environmental indicators. J. Bryol. 2019, 41, 350–360. [Google Scholar] [CrossRef]
- Tusiime, F.M.; Byarujali, S.M.; Bates, J.W. Diversity and distribution of bryophytes in three forest types of Bwindi Impenetrable National Park, Uganda. Afr. J. Ecol. 2007, 45, 79–87. [Google Scholar] [CrossRef]
- Ah-Peng, C.; Wilding, N.; Kluge, J.; Descamps-Julien, B.; Bardat, J.; Chuah-Petiot, M.; Strasberg, D.; Hedderson, T.A. Bryophyte diversity and range size distribution along two altitudinal gradients: Continent vs. island. Acta Oecol. 2012, 42, 58–65. [Google Scholar] [CrossRef]
- Sun, S.Q.; Wu, Y.H.; Wang, G.X.; Zhou, J.; Yu, D.; Bing, H.J.; Luo, J. Bryophyte species richness and composition along an altitudinal gradient in Gongga Mountain, China. PLoS ONE 2013, 8, 1–10. [Google Scholar]
- Rosenzweig, M.L. Species diversity gradients: We know more and less than we thought. J. Mammal. 1992, 73, 715–730. [Google Scholar] [CrossRef]
- Pounds, J.A.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.; Foster, P.N.; Young, B.E. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Sanders, N.J.; Lessard, J.P.; Fitzpatrick, M.C.; Dunn, R.R. Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains. Glob. Ecol. Biogeogr. 2007, 16, 640–649. [Google Scholar] [CrossRef]
- Bryant, J.A.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Cordero, V. Elevation gradients of diversity for rodents and bats in Oaxaca, Mexico. Glob. Ecol. Biogeogr. 2001, 10, 63–76. [Google Scholar] [CrossRef]
- Uchida, M.; Muraoka, H.; Nakatsubo, T.; Bekku, Y.; Ueno, T.; Kanda, H.; Koizumi, H. Net photosynthesis, respiration, and production of the moss Sanionia uncinata on a glacier foreland in the high arctic, Ny-Ålesund, Svalbard. Arct. Antarct. Alp. Res. 2002, 34, 287–292. [Google Scholar] [CrossRef]
- Metcalfe, D.B.; Ahlstrand, J.C.M. Effects of moisture dynamics on bryophyte carbon fluxes in a tropical cloud forest. New Phytol. 2019, 222, 1766–1777. [Google Scholar] [CrossRef] [PubMed]
- Skre, O.; Oechel, W.C. Moss functioning in different taiga ecosystems in interior Alaska: I. Seasonal, phenotypic, and drought effects on photosynthesis and response patterns. Oecologia 1981, 48, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Jónsdóttir, I.S.; Magnússon, B.; Gudmundsson, J.; Elmarsdóttir, Á.; Hjartarson, H. Variable sensitivity of plant communities in Iceland to experimental warming. Glob. Chang. Biol. 2005, 11, 553–563. [Google Scholar] [CrossRef]
- Walker, M.D.; Wahre, C.H.; Hollister, R.D.; Henry, G.H.R.; Ahlquist, L.E.; Alatalo, J.M.; Wookey, P.A. Plant community responses to experimental warming across the tundra biome. Proc. Natl. Acad. Sci. USA 2006, 103, 1342–1346. [Google Scholar] [CrossRef] [Green Version]
- Rincon, E. The effect of herbaceous litter on bryophyte growth. J. Bryol. 1988, 15, 209–217. [Google Scholar] [CrossRef]
- Frahm, J.-P.; Gradstein, S.R. An altitudinal zonation of tropical rain forests using bryophytes. J. Biogeogr. 1991, 18, 669–678. [Google Scholar] [CrossRef]
- Proctor, M.C.F. Physiological ecology. In Bryophyte Biology; Goffinet, B., Shaw, A.J., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 237–268. [Google Scholar]
- Mägdefrau, K. Life-forms of Bryophytes. In Bryophyte Ecology; Smith, A.J.E., Ed.; Springer: Dordrecht, The Netherlands, 1982; pp. 45–58. [Google Scholar]
- Bates, J. Is ‘life-form’ a useful concept in bryophyte ecology? Oikos 1998, 82, 223–237. [Google Scholar] [CrossRef]
- Zhang, L.; Corlett, R. Phytogeography of Hong Kong bryophytes. J. Biogeogr. 2003, 30, 1329–1337. [Google Scholar] [CrossRef]
- Wu, Z.-Y.; Zhou, Z.-K.; Li, D.-Z.; Peng, H.; Sun, H. The areal types of the world families of seed plants. Acta Bot. Yunnanica 2003, 25, 245–257. (In Chinese) [Google Scholar]





| Altitude | AT | DP | RH | RF | pH | GC | CC | WS | VT |
|---|---|---|---|---|---|---|---|---|---|
| 100 | 27.2 a | 23.4 a | 84.8 d | 1838 b | 4.99 a | 89.3 c | 88.8 b | 12.3 b | Dense broad-leaved lowland woodland (Glochidion hongkongense) |
| 300 | 25.4 b | 22.6 b | 88.2 c | 1917 ab | 4.68 b | 94.7 a | 90.0 a | 5.30 d | Dense broad-leaved lowland woodland (Syzgium levinei) |
| 500 | 23.9 c | 22.0 c | 91.8 b | 1992 ab | 4.27 c | 92.0 b | 88.7 c | 7.70 c | Dense broad-leaved low-hill forest (Cinnamomum porrectum) |
| 700 | 23.2 d | 21.9 c | 92.6 ab | 2077 ab | 4.54 b | 53.5 e | 41.9 e | 25.9 a | Grassland with small montane shrub patches (Miscanthus sinensis) |
| 900 | 21.0 e | 19.7 d | 93.4 a | 2271 a | 4.60 b | 68.4 d | 45.0 d | 25.3 a | Grassland with dense montane shrub patches (Miscanthus sinensis) |
| Site | Altitude (m asl) | Species Richness | Cover (%) | Thickness (mm) |
|---|---|---|---|---|
| 1 | 100 | 8.0 ± 1.2 a | 15.0 ± 1.0 b | 3.23 ± 0.3 b |
| 2 | 300 | 7.7 ± 1.2 a | 10.4 ± 0.2 c,d | 3.23 ± 0.2 b |
| 3 | 500 | 7.0 ± 0.6 a | 12.3 ± 1.8 b,c | 3.47 ± 0.3 b |
| 4 | 700 | 5.0 ± 1.2 a | 7.2 ± 0.1 d | 5.07 ± 0.5 b |
| 5 | 900 | 7.7 ± 1.2 a | 23.8 ± 0.6 a | 8.60 ± 0.7 a |
| General Patterns | Moss Species (% Total) | Patterns in Appendix C Table A3 |
|---|---|---|
| Tropical | 33 (64.7%) | 2, 3, 4, 5, 7 |
| East Asian | 24 (47.1%) | 5, 6, 7, 9 |
| Temperate | 8 (15.7%) | 8 |
| Total | 51 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hao, J.; Chu, L.M. Vegetation Types Attributed to Deforestation and Secondary Succession Drive the Elevational Changes in Diversity and Distribution of Terrestrial Mosses in a Tropical Mountain Forest in Southern China. Forests 2021, 12, 961. https://doi.org/10.3390/f12080961
Hao J, Chu LM. Vegetation Types Attributed to Deforestation and Secondary Succession Drive the Elevational Changes in Diversity and Distribution of Terrestrial Mosses in a Tropical Mountain Forest in Southern China. Forests. 2021; 12(8):961. https://doi.org/10.3390/f12080961
Chicago/Turabian StyleHao, Jiewei, and L. M. Chu. 2021. "Vegetation Types Attributed to Deforestation and Secondary Succession Drive the Elevational Changes in Diversity and Distribution of Terrestrial Mosses in a Tropical Mountain Forest in Southern China" Forests 12, no. 8: 961. https://doi.org/10.3390/f12080961