1. Introduction
Enteromorpha clathrata is an edible alga that has been widely consumed in Asian countries [
1,
2,
3,
4]. Previous studies have indicated that
Enteromorpha clathrata polysaccharide (ECP) could be used as a novel drug candidate to treat obesity, constipation and hyperlipidemia [
3,
5,
6]. However, what effect it has on inflammatory bowel diseases has not been investigated. Ulcerative colitis and Crohn’s disease are two types of typical inflammatory bowel diseases, and preceding results have illustrated a causal role of gut dysbiosis in the development of ulcerative colitis [
7,
8,
9,
10]. ECP is not absorbed after oral intake and therefore holds great potential for the treatment of dysbiosis-associated intestinal diseases by targeting gut microbiota [
1,
2,
4].
Our previous results suggested that dietary intake of ECP could modulate the composition of the gut microbiota and increase the abundances of beneficial microbes in the gut, including
Akkermansia muciniphila,
Eubacterium xylanophilum,
Bifidobacterium spp. and
Lactobacillus spp. [
1,
5]. Similarly, Ren et al. found that ECP could ameliorate loperamide-induced gastrointestinal disorders by increasing the populations of probiotic bacteria such as
Prevotella spp. and
Bacteroides spp. in the colon [
6]. Taken together, the prebiotic effect of ECP prompted us to investigate whether it also has any therapeutic effects on inflammatory bowel diseases.
In the present study, using a mouse model of dextran sulfate sodium (DSS)-induced ulcerative colitis, we illustrate for the first time a beneficial effect of ECP on ulcerative colitis and provide a possible basis of understanding its therapeutic mechanisms from the perspective of symbiotic gut bacteria Parabacteroides distasonis. Our study paves the way for the development of ECP as a new drug candidate for the treatment of inflammatory bowel diseases.
2. Results
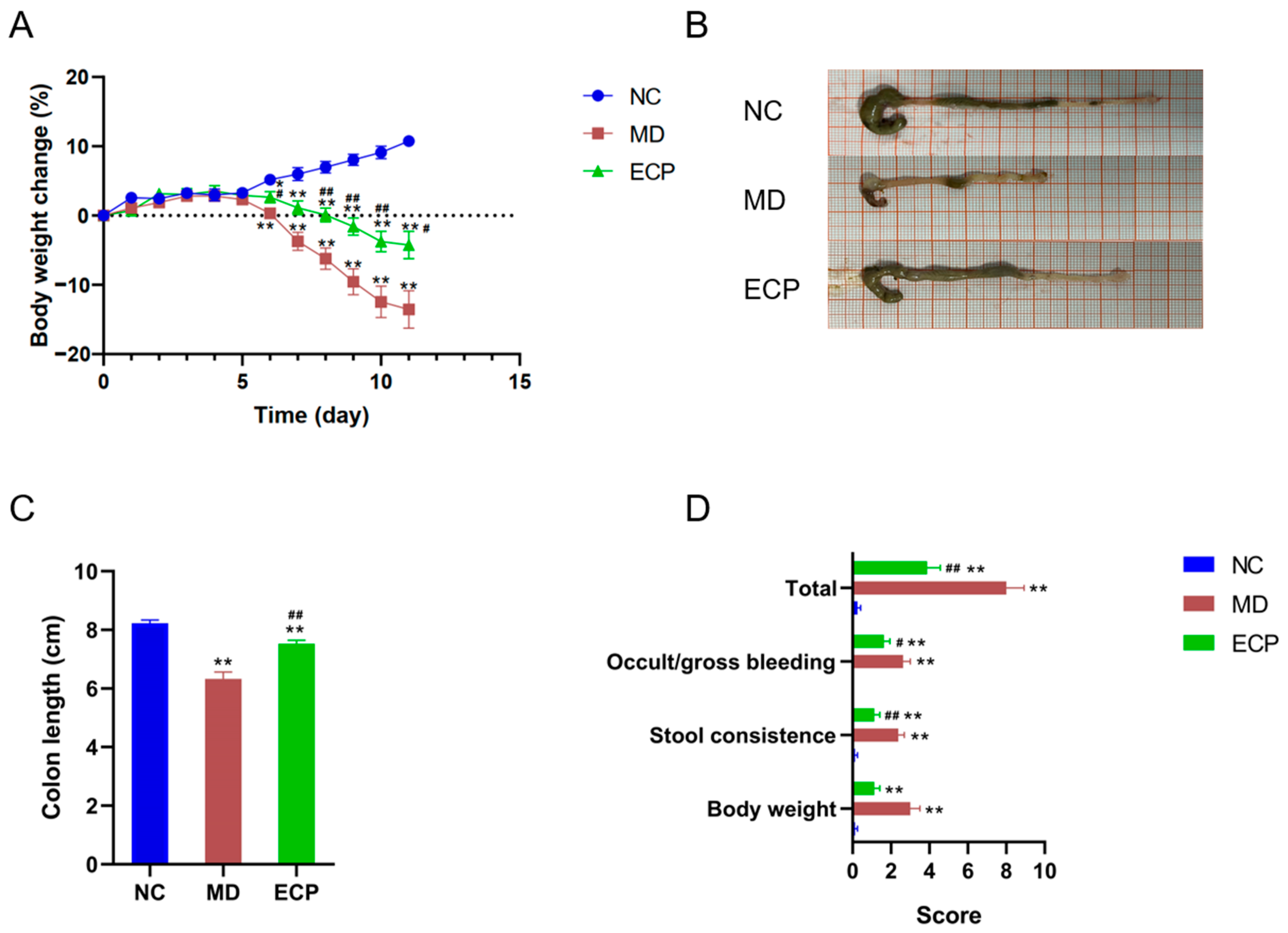
2.1. Dietary ECP Improved Ulcerative Colitis and Ameliorated Mucosal Damage in DSS-Fed Mice
We explored the therapeutic effect of ECP on ulcerative colitis using a well-established mouse model [
11,
12]. DSS-induced ulcerative colitis in mice is associated with decreased body weight and shortening of the colon length. Interestingly, dietary intake of ECP (100 mg/kg) considerably retarded the body weight loss in DSS-treated mice (
Figure 1A). Additionally, the shortening of colon length in diseased mice was also profoundly suppressed by the treatment of ECP (
Figure 1B,C). Furthermore, oral administration of ECP also significantly reduced the incidences of colonic bleeding and improved stool consistency in diseased mice (
Figure 1D).
Previous studies have indicated that DSS-induced ulcerative colitis is correlated with mucosal damage in the colon [
11,
12,
13]. In line with previous results, DSS disrupted the intestinal epithelial layer and inhibited the biosynthesis of colonic mucin
O-glycans (
Figure 2A,B). However, intriguingly, dietary intake of ECP reversed the toxic effect of DSS and attenuated the mucosal damage in the colon of diseased mice, as indicated by histochemical staining (
Figure 2A,B) and histopathological colitis score analysis (
Figure 2C).
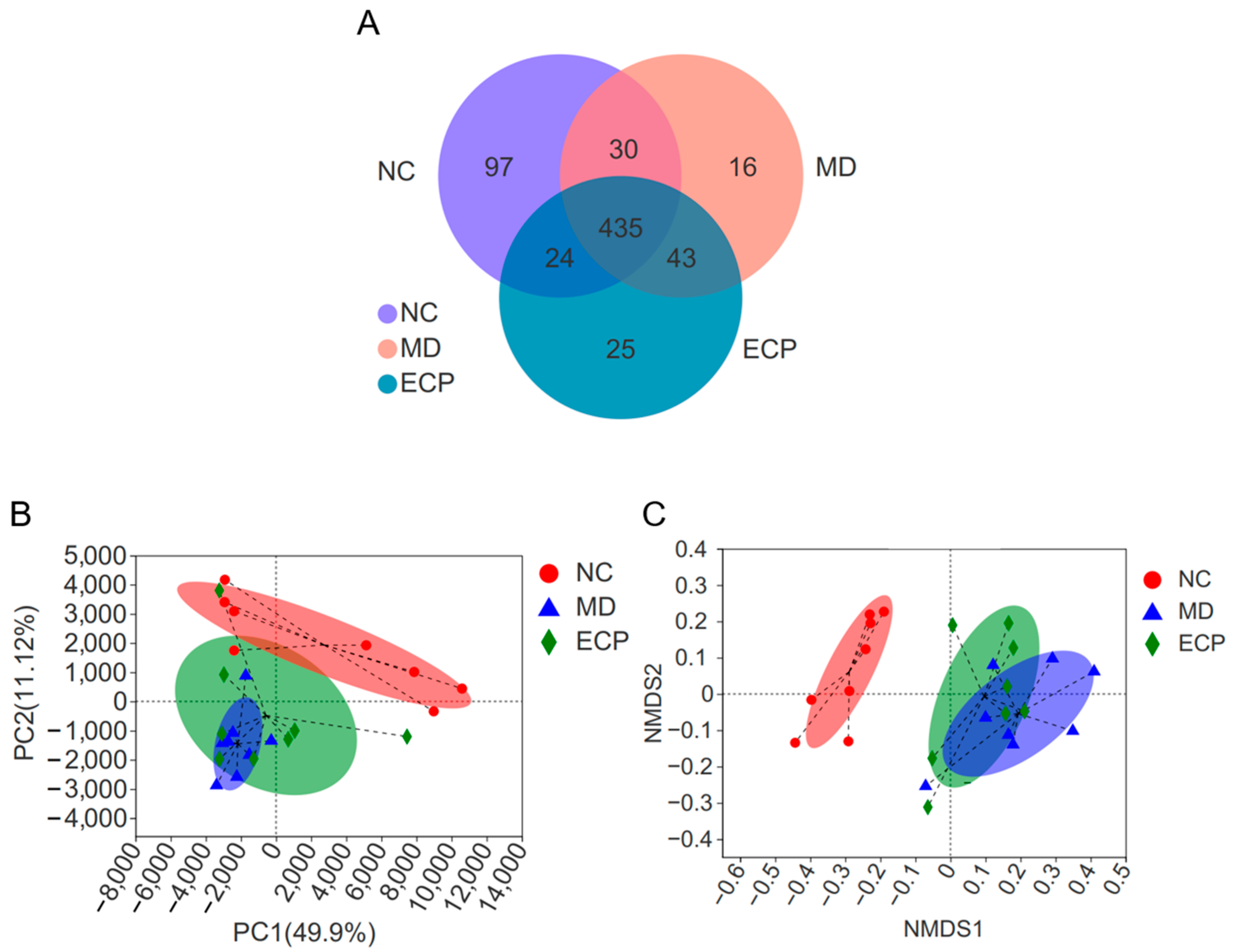
2.2. Dietary ECP Changed the Overall Structure of the Gut Microbiota in DSS-Fed mice
Previous studies have indicated that ECP is not absorbed after oral intake [
1,
2,
4]. Therefore, when reaching the colon, it would be fermented by the intestinal microbes [
1,
4,
5]. In this regard, we next sought to explore the beneficial effects of ECP on ulcerative colitis from the perspective of gut microbiota. Expectedly, dietary ECP significantly changed the overall structure of the gut microbiota in DSS-fed mice. Venn diagram analysis suggested that 49 and 46 operational taxonomic units (OTUs) were separately identified in ECP-treated mice and diseased mice (
Figure 3A). Additionally, as indicated by principal components analysis (PCA) and non-metric multidimensional scaling (NMDS) score plot analyses, dietary ECP induced a remarkable shift of gut microbiota structure towards that of healthy mice (
Figure 3B,C).
2.3. Dietary ECP Modulated the Composition of the Gut Microbiota and Increased the Abundance of Parabacteroides spp. in DSS-Fed Mice
The mice gut microbes were dominated by Bacteroidota, Actinobacteriota and Firmicutes at the phylum level and
Dubosiella spp.,
Bacteroides spp.,
Lactobacillus spp.,
Alistipes spp.,
Parabacteroides spp. and
Blautia spp. at the genus level (
Figure 4). Heatmap analysis of the gut microbiota showed that intake of ECP could potentially affect the composition of the gut microbiota (
Figure 4).
To identify the key bacteria that were changed by ECP treatment, we performed LEfSe analysis. NC mice were characterized by higher abundances of
Lachnospiraceae NK4A136 spp.,
Lactobacillus johnsonii and
Muribaculaceae compared to DSS-fed mice (
Figure 5A). In line with the preceding results [
13], DSS-induced ulcerative colitis was associated with increased amounts of mucolytic bacteria, including
Akkermansia muciniphila,
Bacteroides vulgatus,
Bacteroides acidifaciens and
Romboutsia ilealis (
Figure 5A). Interestingly, ECP treatment remarkably decreased the amount of mucolytic bacteria, including
Akkermansia muciniphila and
Bacteroides thetaiotaomicron and promoted the growth of potential beneficial bacteria such as
Parabacteroides spp. and
Alistipes spp. (
Figure 5B).
We next tried to isolate the probable colitis-protective bacteria in the genus of
Parabacteroides spp. and
Alistipes spp. from the colon contents of ECP-treated mice. However, we were not able to obtain the targeted bacteria. For this reason, we then turned to
Parabacteroides distasonis F1-28, a bacterium that has been previoulsy isolated from the feces of a healthy individual in our lab.
P. distasonis was chosen because previous studies have indicated that it could alleviates 2,4,6-trinitrobenzenesulfonic acid (TNBS)-induced colitis [
14,
15].
P. distasonis has been proposed as a next-generation probitic bacterium [
14], and we hypothesized that ECP might be fermentated by
P. distasonis F1-28, and
P. distasonis F1-28 might also be protective against DSS-induced ulcerative colitis.
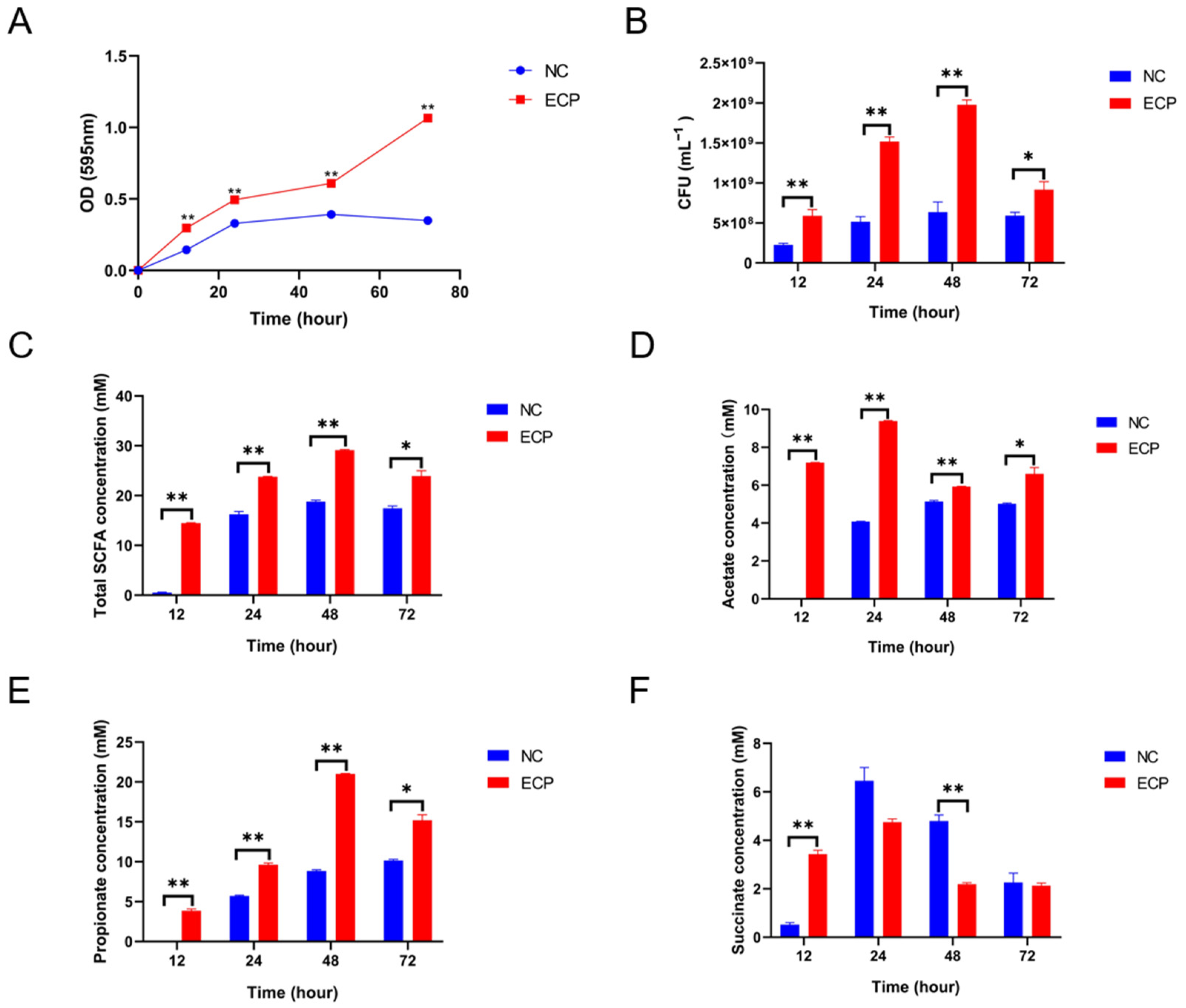
2.4. ECP Promoted the Growth of P. distasonis F1-28 In Vitro and Increased the Production of Short-Chain Fatty Acids
To test the aforementioned hypothesis, ECP was added to the culture medium of
P. distasonis F1-28. Interestingly, ECP significantly promoted the growth of
P. distasonis F1-28 in vitro (
Figure 6A,B). Short-chain fatty acids (SCFAs), including acetate, propionate and succinate, are major fermentation products of
P. distasonis [
14]. In the present study, ECP considerably increased the productions of total SCFAs, acetate, propionate and succinate at different fermentation periods (
Figure 6C–F). Altogether, our study indicates that ECP could be utilized and fermented by
P. distasonis in the gut, and this could help to explain why the abundance of
Parabacteroides spp. was increased in ECP-treated mice.
2.5. P. distasonis F1-28 Alleviated Ulcerative Colitis and Attenuated Mucosal Damage in DSS-Fed Mice
In light of the fact that ECP promotes the growth of
P. distasonis F1-28, we next sought to investigate whether
P. distasonis F1-28 has any therapeutic effects on DSS-induced colitis. Interestingly, we found that oral administration of live
P. distasonis F1-28 could alleviate ulcerative colitis in DSS-fed mice. Specifically, intake of
P. distasonis F1-28 retarded the body weight loss, reduced the contraction of the colon length, decreased the incidences of bleeding and improved stool consistency in diseased mice (
Figure 7A–D).
Histochemical staining and histopathological colitis score analysis suggested that
P. distasonis F1-28 could also attenuate DSS-induced mucosal damage in the colon (
Figure 8A–C). Altogether, our study corroborates previous results showing an anti-inflammatory effect of
P. distasonis on TNBS-induced colitis [
15]. ECP could promote the growth of
P. distasonis F1-28, and this could help to understand the therapeutic effect of ECP on chemical-induced colitis in mice.
3. Discussion
Enteromorpha clathrata is a well-known edible alga that has been traditionally consumed in Asian countries, including China, Korea and Japan [
1,
2,
3,
4].
Enteromorpha clathrata polysaccharide (ECP) has been proposed as a novel drug candidate to treat obesity, constipation and hyperlipidemia [
3,
5,
6]. However, what effect it has on inflammatory bowel diseases has not been investigated. Our study demonstrates for the first time a beneficial effect of ECP on ulcerative colitis and provides a possible basis for understanding its therapeutic mechanisms from the perspective of symbiotic gut bacteria
P. distasonis. Our study paves the way for the development of ECP as a new marine drug candidate for the treatment of inflammatory bowel diseases such as ulcerative colitis and Crohn’s disease.
Our study has two limitations. First, due to the experimental design, we were not able to determine whether the beneficial effects of ECP on gut microbiota was a cause or consequence of the attenuated colitis. Therefore, future studies using fecal microbiota transplantation and germ-free mice are encouraged to further explore this question. Second, our study could only provide a possible basis for understanding the therapeutic mechanisms of ECP from the perspective of symbiotic gut bacteria P. distasonis. This is partially because we failed to isolate any P. distasonis strains from the ECP-treated mice. It is hypothesized that ECP could be fermented in vivo by specific P. distasonis bacterium. However, more detailed studies are needed to explore this issue.
In addition to
P. distasonis,
P. goldsteinii is also recognized as a potential next-generation probiotic candidate due to its protective effect on inflammation [
16]. Recently,
P. goldsteinii was found to attenuate ulcerative colitis in mice treated with DSS [
17]. Similarly,
Alistipes finegoldii was also demonstrated to have a protective effect on DSS-induced colitis [
18]. Our study indicated that ECP could increase the abundances of
Alistipes spp. and
Parabacteroides spp. in DSS-fed mice during alleviation of colitis symptoms. Although we focused on
P. distasonis in our present study, we do not rule out the possibility that the anti-colitis effect of ECP may be mediated by other probiotic bacteria in the gut, such as
P. goldsteinii and
A. finegoldii.
Intestinal bile acids play a pivotal role in the pathogenesis of inflammatory bowel diseases [
19].
P. distasonis has been suggested to possess a wide range of bile acid conversion functions [
20]. Previous studies have indicated that secondary bile acids, including ursodeoxycholic acid and lithocholic acid, could protect against intestinal inflammation by inhibiting the apoptosis of epithelial cells and restoring the integrity of the mucosal layer [
21]. Further investigations are therefore needed to explore the effect of ECP on the metabolism of bile acids during alleviation of ulcerative colitis.
4. Materials and Methods
4.1. Chemicals and Reagents
The
Enteromorpha clathrata polysaccharide (ECP) was used and prepared as previously described [
1,
5]. Tryptone, peptone, yeast extract and Tween 80 were obtained from Sigma (Shanghai, China). Hemin and L-cysteine hydrochloride were purchased from Sangon Biotech (Shanghai, China). All other chemicals of analytical grade were acquired from Sinopharm Chemical (Shanghai, China).
4.2. Animals and Treatment
The animal experiments in the present study were approved and supported by the Ethical Committee of Ocean University of China, School of Medicine and Pharmacy (Permission No. OUC-2021-0301-02) and complied with the Guide for the Care and Use of Laboratory Animals (National Academies Press, 8th edition, 2011). All the animals were obtained from Beijing Vital River Laboratory Animal Technology Co. Ltd. (Beijing, China) (Certificate No. SCXK (Jing) 2016-0011).
In animal experiment 1, a total of 24 eight-week-old male C57BL/6J specific pathogen-free (SPF) mice were used to test the anti-colitis effect of ECP. The mice were divided into three groups: the normal control group (NC, n = 8), the model group (MD, n = 8) and the ECP treatment group (ECP, n = 8). ECP was dissolved in phosphate-buffered saline (PBS) and was given at a dosage of 100 mg/kg/day by gavage. Mice in MD group and ECP group were given 2.0% (w/v) DSS (MP Biomedicals LLC, Solon, USA) in the daily drinking water for 8 consecutive days. All mice were humanely sacrificed on the eleventh day and the colon and cecum were collected for further experiment.
In animal experiment 2, a total of 24 eight-week-old male C57BL/6J specific pathogen-free (SPF) mice were used to test the anti-colitis effect of P. distasonis F1-28. The mice were divided into three groups: the normal control group (NC, n = 8), the model group (MD, n = 8) and the P. distasonis F1-28 treatment group (PD, n = 8). P. distasonis F1-28 was dissolved in phosphate-buffered saline (PBS) and was given at a dosage of about 2.0 × 107 colony-forming units (CFUs)/day/mouse by gavage. Mice in MD group and ECP group were given 2.0% (w/v) DSS in the daily drinking water for 7 consecutive days. All mice were humanely sacrificed on the ninth day and the colon and cecum were collected for further experiment.
The body weight and the stool morphology of the mice were monitored every day. H&E staining and Alcian blue staining of the colon tissues were performed as previously described [
13,
22,
23]. The symptom score and histopathological colon score analyses were performed as described elsewhere [
13,
24,
25].
4.3. In Vitro Fermentation of P. distasonis F1-28
The VI medium was applied to investigate the utilization and fermentation of ECP by
P. distasonis F1-28 as previously described [
16,
17]. In this in vitro fermentation experiment,
P. distasonis F1-28 was grown in the basic VI medium that contained no extra carbon source (NC group) and the basic medium that contained ECP as the primary carbon source (ECP group). The CFUs of
P. distasonis F1-28 was checked at different time points. The SCFAs levels in the two media were analyzed using the methods described elsewhere [
26,
27].
4.4. High-Throughput Sequencing and Bioinformatic Analyses
The metagenomic DNA of the gut microbiota were extracted using the commercial Qiagen QIAamp DNA Stool Mini Kit (Hilden, Germany) from the cecum samples. The 16S V3-V4 hypervariable gene regions were specifically amplified using the well-established universal primers 338F (ACTCCTACGGGAGGAGCAG) and 806R(GGACTACHVGGGTWTCTAAT). The amplicons were quality-checked and sequenced on an Illumina PE300 platform from Majorbio Bio-pharm Biotechnology Co., Ltd. (Shanghai, China). Bioinformatic analyses of the sequencing data (Venn diagram, PCA score plot, NMDS score plot and heatmap analyses) were conducted using the online Majorbio Cloud Platform (
www.majorbio.com (accessed on 27 July 2022)). The LEfSe analysis was performed as previously described [
26,
27]. Only bacterial taxa with an LDA score of above 4.0 and 3.0 were listed.
4.5. Statistical Analyses
Data were expressed as mean ± SEM. Statistical analyses were performed using Student t-test and ANOVA with post-hoc Tukey’s tests (GraphPad Prism for Windows 8.0; GraphPad Software Inc, San Diego, CA, USA). The results were considered statistically significant at p < 0.05. * p < 0.05 versus NC group; ** p < 0.01 versus NC group; # p < 0.05 versus MD group; ## p < 0.01 versus MD group.
5. Conclusions
In conclusion, dietary ECP improved ulcerative colitis and ameliorated mucosal damage in DSS-fed mice. ECP changed the structure of the gut microbiota in diseased mice by increasing the abundance of probiotic bacteria Parabacteroides spp. In vitro, ECP promoted the growth of anti-colitis bacterium P. distasonis F1-28 and increased its production of SCFAs. Our study demonstrates a therapeutic effect of ECP on DSS-induced ulcerative colitis and provides a possible basis for understanding its mechanisms from the perspective of symbiotic gut bacteria P. distasonis.
Author Contributions
Conceptualization, Q.S. and G.Y.; methodology, Q.S., A.Z. and P.G.; software, Q.S.; validation, Q.S.; formal analysis, Q.S. and M.M.; investigation, M.M., T.F., Y.W., A.Z. and P.G.; resources, Q.S., A.Z. and P.G.; data curation, M.M., T.F. and Y.W.; writing—original draft preparation, Q.S. and M.M.; writing—review and editing, Q.S.; visualization, Q.S. and M.M.; supervision, Q.S. and G.Y.; project administration, Q.S. and G.Y.; funding acquisition, Q.S. and G.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (81991522, 32101032), Shandong Major Science and Technology Project (2021ZDSYS22), Taishan Scholar Climbing Project (TSPD20210304) and Natural Science Foundation of Shandong Province (ZR2021QC110).
Institutional Review Board Statement
The animal study protocol was approved by the Institutional Review Board (or Ethics Committee) of Ocean University of China, School of Medicine and Pharmacy (Permission No. OUC-2021-0301-02) (protocol code OUC-2021-0302 and date of approval 1 March 2021).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Shang, Q.; Wang, Y.; Pan, L.; Niu, Q.; Li, C.; Jiang, H.; Cai, C.; Hao, J.; Li, G.; Yu, G. Dietary polysaccharide from Enteromorpha clathrata modulates gut microbiota and promotes the growth of Akkermansia muciniphila, Bifidobacterium spp. and Lactobacillus spp. Mar. Drugs 2018, 16, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheong, K.L.; Yu, B.; Chen, J.; Zhong, S. A comprehensive review of the cardioprotective effect of marine algae polysaccharide on the gut microbiota. Foods 2022, 11, 3550. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Gao, H.; Wang, S.; Wen, S.; Qin, S. Hypolipidemic and antioxidant properties of a polysaccharide fraction from Enteromorpha prolifera. Int. J. Biol. Macromol. 2013, 58, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.; Jiang, H.; Cai, C.; Hao, J.; Li, G.; Yu, G. Gut microbiota fermentation of marine polysaccharides and its effects on intestinal ecology: An overview. Carbohydr. Polym. 2018, 179, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Zhao, Y.; Zhou, C.; Zhao, Q.; Zhong, H.; Zhu, X.; Fu, T.; Pan, L.; Shang, Q.; Yu, G. Dietary polysaccharide from Enteromorpha clathrata attenuates obesity and increases the intestinal abundance of butyrate-producing bacterium, Eubacterium xylanophilum, in mice fed a high-fat diet. Polymers 2021, 13, 3286. [Google Scholar] [CrossRef]
- Ren, X.; Liu, L.; Gamallat, Y.; Zhang, B.; Xin, Y. Enteromorpha and polysaccharides from Enteromorpha ameliorate loperamide-induced constipation in mice. Biomed. Pharmacother. 2017, 96, 1075–1081. [Google Scholar] [CrossRef]
- GBD 2017 Inflammatory Bowel Disease Collaborators. The global, regional, and national burden of inflammatory bowel disease in 195 countries and territories, 1990-2017: A systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol. Hepatol. 2020, 5, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Kudelka, M.R.; Stowell, S.R.; Cummings, R.D.; Neish, A.S. Intestinal epithelial glycosylation in homeostasis and gut microbiota interactions in IBD. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 597–617. [Google Scholar] [CrossRef]
- Lavelle, A.; Sokol, H. Gut microbiota-derived metabolites as key actors in inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 223–237. [Google Scholar] [CrossRef]
- Schirmer, M.; Garner, A.; Vlamakis, H.; Xavier, R.J. Microbial genes and pathways in inflammatory bowel disease. Nat. Rev. Microbiol. 2019, 17, 497–511. [Google Scholar] [CrossRef]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran sulfate sodium (DSS)-induced colitis in mice. Curr. Protoc. Immunol. 2014, 104, 15.25.1–15.25.14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirtz, S.; Popp, V.; Kindermann, M.; Gerlach, K.; Weigmann, B.; Fichtner-Feigl, S.; Neurath, M.F. Chemically induced mouse models of acute and chronic intestinal inflammation. Nat. Protoc. 2017, 12, 1295–1309. [Google Scholar] [CrossRef]
- Pan, L.; Fu, T.; Cheng, H.; Mi, J.; Shang, Q.; Yu, G. Polysaccharide from edible alga Gloiopeltis furcata attenuates intestinal mucosal damage by therapeutically remodeling the interactions between gut microbiota and mucin O-glycans. Carbohydr. Polym. 2022, 278, 118921. [Google Scholar] [CrossRef]
- Ezeji, J.C.; Sarikonda, D.K.; Hopperton, A.; Erkkila, H.L.; Cohen, D.E.; Martinez, S.P.; Cominelli, F.; Kuwahara, T.; Dichosa, A.E.K.; Good, C.E.; et al. Parabacteroides distasonis: Intriguing aerotolerant gut anaerobe with emerging antimicrobial resistance and pathogenic and probiotic roles in human health. Gut Microbes 2021, 13, 1922241. [Google Scholar] [CrossRef] [PubMed]
- Cuffaro, B.; Assohoun, A.L.W.; Boutillier, D.; Súkeníková, L.; Desramaut, J.; Boudebbouze, S.; Salomé-Desnoulez, S.; Hrdý, J.; Waligora-Dupriet, A.-J.; Maguin, E.; et al. In vitro characterization of gut microbiota-derived commensal strains: Selection of Parabacteroides distasonis strains alleviating TNBS-induced colitis in mice. Cells 2020, 9, 2104. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Zhang, L.; Wang, X.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Roles of intestinal Parabacteroides in human health and diseases. FEMS Microbiol. Lett. 2022, 369, fnac072. [Google Scholar] [CrossRef]
- Gerkins, C.; Oliero, M.; Hajjar, R.; Rendos, H.V.; Fragoso, G.; Calvé, A.; Diop, K.; Routy, B.; Santos, M. The modulation of intestinal inflammation by parabacteroides goldsteinii in dextran sodium sulfate induced colitis in mice. Gut 2022, 71, A60. [Google Scholar]
- Dziarski, R.; Park, S.Y.; Kashyap, D.R.; Dowd, S.E.; Gupta, D. Pglyrp-regulated gut microflora Prevotella falsenii, Parabacteroides distasonis and Bacteroides eggerthii enhance and Alistipes finegoldii attenuates colitis in mice. PLoS ONE 2016, 11, e0146162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiratterra, E.; Franco, P.; Porru, E.; Katsanos, K.H.; Christodoulou, D.K.; Roda, G. Role of bile acids in inflammatory bowel disease. Ann. Gastroenterol. 2018, 31, 266–272. [Google Scholar] [PubMed]
- Wang, K.; Liao, M.; Zhou, N.; Bao, L.; Ma, K.; Zheng, Z.; Wang, Y.; Liu, C.; Wang, W.; Wang, J.; et al. Parabacteroides distasonis Alleviates Obesity and Metabolic Dysfunctions via Production of Succinate and Secondary Bile Acids. Cell Rep. 2019, 26, 222–235.e5. [Google Scholar] [CrossRef] [Green Version]
- Lajczak-McGinley, N.K.; Porru, E.; Fallon, C.M.; Smyth, J.; Curley, C.; McCarron, P.A.; Tambuwala, M.M.; Roda, A.; Keely, S.J. The secondary bile acids, ursodeoxycholic acid and lithocholic acid, protect against intestinal inflammation by inhibition of epithelial apoptosis. Physiol. Rep. 2020, 8, e14456. [Google Scholar] [CrossRef]
- Yang, D.; Jacobson, A.; Meerschaert, K.A.; Sifakis, J.J.; Wu, M.; Chen, X.; Yang, T.; Zhou, Y.; Anekal, P.V.; Rucker, R.A.; et al. Nociceptor neurons direct goblet cells via a CGRP-RAMP1 axis to drive mucus production and gut barrier protection. Cell 2022, 185, 4190–4205.e5. [Google Scholar] [CrossRef] [PubMed]
- Tawfiq, R.A.; Nassar, N.N.; Hammam, O.A.; Allam, R.M.; Elmazar, M.M.; Abdallah, D.M.; Attia, Y.M. Obeticholic acid orchestrates the crosstalk between ileal autophagy and tight junctions in non-alcoholic steatohepatitis: Role of TLR4/TGF-β1 axis. Chem. Biol. Interact. 2022, 361, 109953. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, M.; Chen, Q.; Li, X.; Chen, L.; Dong, Z.; Zhu, W.; Yang, Y.; Liu, Z.; Chen, Q. Programmable probiotics modulate inflammation and gut microbiota for inflammatory bowel disease treatment after effective oral delivery. Nat. Commun. 2022, 13, 3432. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, D.; Liang, Y.; Zhang, Z.; Guo, J.; Chen, Y.; Yan, Y.; Liu, H.; Lei, L.; Wang, Z.; et al. Licorice-Yuanhua Herbal Pair Induces Ileum Injuries Through Weakening Epithelial and Mucous Barrier Functions: Saponins, Flavonoids, and Di-Terpenes All Involved. Front. Pharmacol. 2020, 11, 869. [Google Scholar] [CrossRef]
- Fu, T.; Pan, L.; Shang, Q.; Yu, G. Fermentation of alginate and its derivatives by different enterotypes of human gut microbiota: Towards personalized nutrition using enterotype-specific dietary fibers. Int. J. Biol. Macromol. 2021, 183, 1649–1659. [Google Scholar] [CrossRef]
- Fu, T.; Zhou, L.; Fu, Z.; Zhang, B.; Li, Q.; Pan, L.; Zhou, C.; Zhao, Q.; Shang, Q.; Yu, G. Enterotype-specific effect of human gut microbiota on the fermentation of marine algae oligosaccharides: A preliminary proof-of-concept in vitro study. Polymers 2022, 14, 770. [Google Scholar] [CrossRef]
| Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}







