

The First Report of Holothuria (Thymiosycia) impatiens (Forsskål, 1775), (Holothuroidea: Holothuriidae) from Tunisia (Mediterranean Sea): Taxonomic, Morphological, and Molecular Data Compilation
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Morphological Identification
Age-Related Variation of Ossicles
2.3. Genetic Study
DNA Extraction and Amplification
2.4. PCA and Biplot Analysis
3. Results
3.1. Morphological Identification
3.1.1. Special Observation
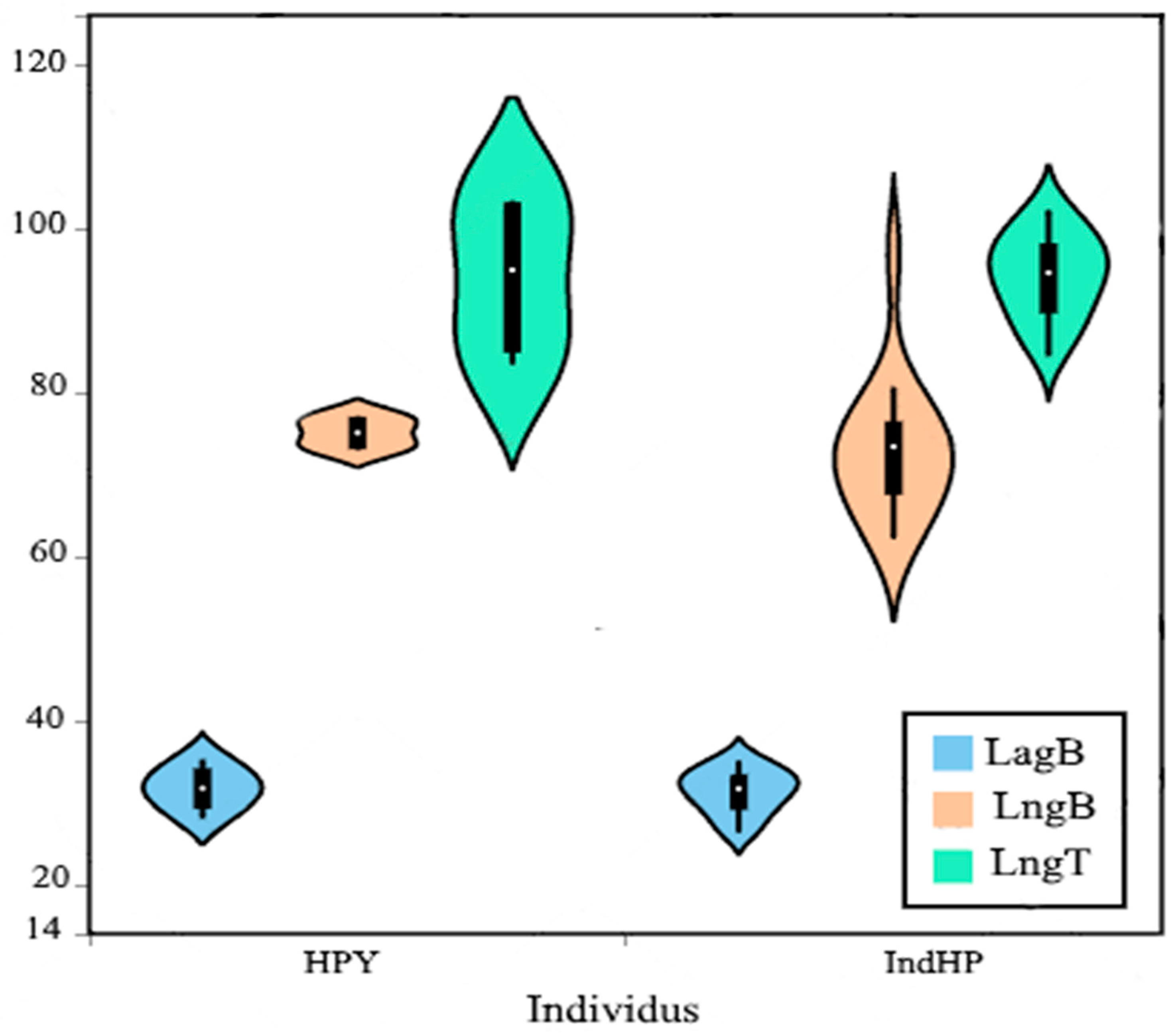
3.1.2. Ossicles Morphology
3.2. Genetic Study (Barcoding of the COI Marker)
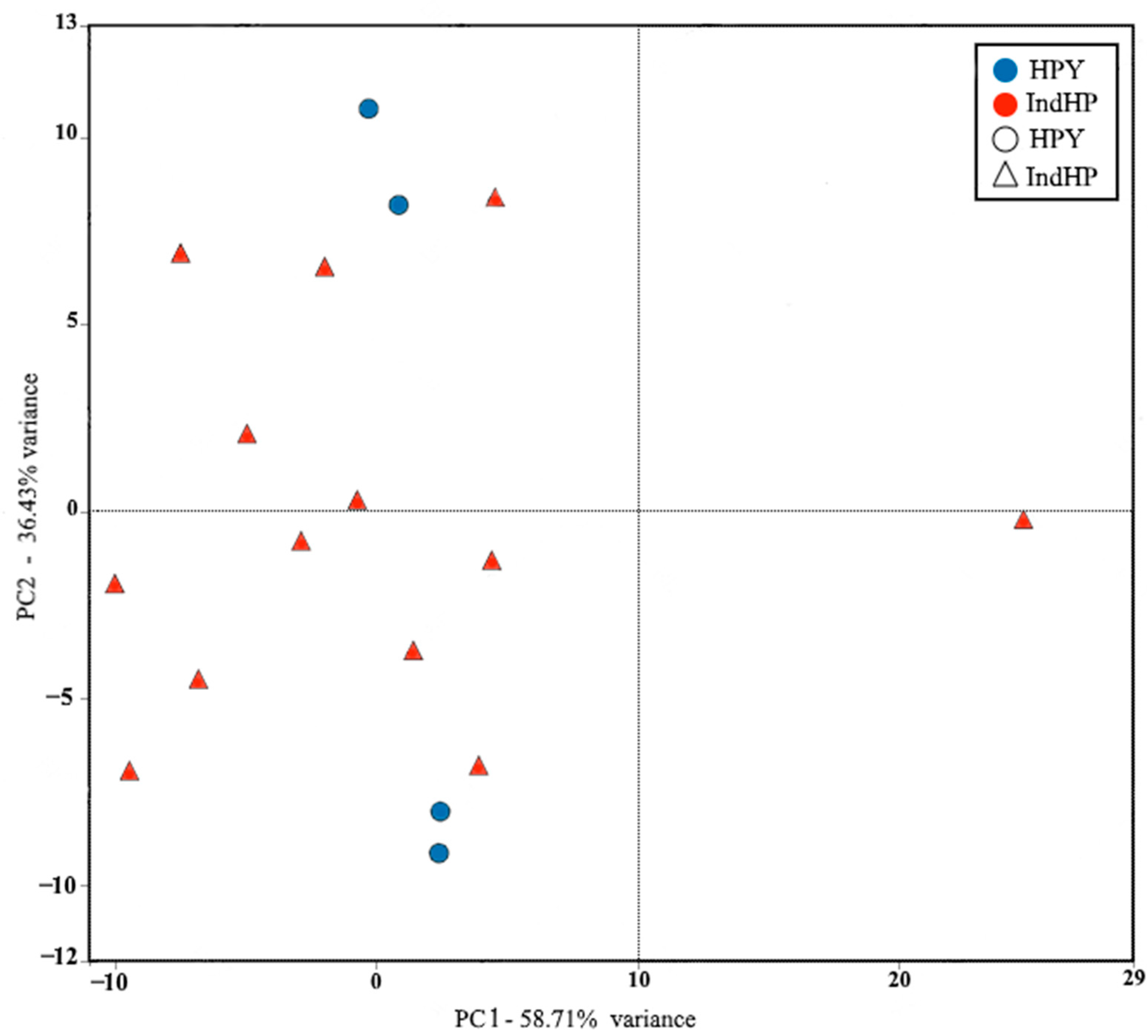
3.3. Principal Component Analysis of the Ossicles
Biplot Analysis of Ossicles
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- O’Loughlin, P.M.; Paulay, G.; Vandenspiegel, D.; Samyn, Y. New Holothuria species from Australia (Echinodermata: Holothuroidea: Holothuriidae), with comments on the origin of deep and cool holothuriids. Mem. Mus. Vic. 2007, 64, 35–52. [Google Scholar] [CrossRef] [Green Version]
- Pérez, G.H.B. Sistemática y Filogeografía De Las Especies Del Subgénero Holothuria (Echinodermata: Holothuriidae: Holothuria) de la Región Atlanto-Mediterránea. Ph.D. Thesis, Universidad de Murcia, Murcia, Spain, 2010. [Google Scholar]
- MacTavish, T.; Stenton-Dozey, J.; Vopel, K.; Savage, C. Deposit-Feeding Sea Cucumbers Enhance Mineralization and Nutrient Cycling in Organically-Enriched Coastal Sediments. PLoS ONE 2012, 7, e50031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, S.C.; Flemming, J.M.; Watson, R.; Lotze, H.K. Serial exploitation of global sea cucumber fisheries. Fish Fish. 2011, 12, 317–339. [Google Scholar] [CrossRef]
- Siegenthaler, A.; Cánovas, F.; González-Wangüemert, M. Spatial distribution patterns and movements of Holothuria arguinensis in the Ria Formosa (Portugal). J. Sea Res. 2015, 102, 33–40. [Google Scholar] [CrossRef] [Green Version]
- González-Wangüemert, M.; Borrero-Pérez, G. A new record of Holothuria arguinensis colonizing the Mediterranean Sea. Mar. Biodivers. Rec. 2012, 5, e105. [Google Scholar] [CrossRef] [Green Version]
- González-Wangüemert, M.; Aydin, M.; Conand, C. Assessment of sea cucumber populations from the Aegean Sea (Turkey): First insights to sustainable management of new fisheries. Ocean Coast. Manag. 2014, 92, 87–94. [Google Scholar] [CrossRef]
- González-Wangüemert, M.; Valente, S.; Henriques, F.; Domínguez-Godino, J.A.; Serrão, E.A. Setting preliminary biometric baselines for new target sea cucumbers species of the NE Atlantic and Mediterranean fisheries. Fish. Res. 2016, 179, 57–66. [Google Scholar] [CrossRef]
- Kœhler, R. Faune de France: Echinodermes. 1921. Available online: https://doris.ffessm.fr/Bibliographie/FAUNE-DE-FRANCE-ECHINODERMES-Koehler-R (accessed on 4 February 2023).
- Boudouresque, C.F. Critères de sélection et projet de liste des espèces en danger ou menacées. In Réunion D’experts sur Les espèces Menacées en Méditerranée; UNEP (OCA)/MED WG: Athens, Greece, 1995; Volume 100. [Google Scholar]
- Mustapha, K.B.; Hattour, A.; M’hetli, M. Etat de la bionomie benthique des étages infra et circalittoral du Golfe de Gabes. INSTM Bull. Mar. Freshw. Sci. 1999, 26, 5–48. [Google Scholar]
- Honey-Escandón, M.; Laguarda-Figueras, A.; Solís-Marín, F.A. Molecular phylogeny of the subgenus Holothuria (Selenkothuria) Deichmann, 1958 (Holothuroidea: Aspidochirotida). Zool. J. Linn. Soc. 2012, 165, 109–120. [Google Scholar] [CrossRef] [Green Version]
- Rowe, F.W.E. A review of the family Holothuriidae (Holothurioidea: Aspidochirotida). Bull. Br. Mus. (Nat. Hist.) Zool. 1969, 18, 117–170. [Google Scholar] [CrossRef]
- Samyn, Y.; Vandenspiegel, D. <strong>Sublittoral and bathyal sea cucumbers (Echinodermata: Holothuroidea) from the Northern Mozambique Channel with description of six new species</strong>. Zootaxa 2016, 4196, 451–497. [Google Scholar] [CrossRef]
- Hedingia Mediterranea (Bartolini Baldelli, 1914) Tortonese. 1965. Available online: https://www.gbif.org/species/4342673 (accessed on 4 February 2023).
- Borrero-Pérez, G.H.; Pérez-Ruzafa, A.; Marcos, C.; González-Wangüemert, M. The taxonomic status of some Atlanto-Mediterranean species in the subgenus Holothuria (Echinodermata: Holothuroidea: Holothuriidae) based on molecular evidence. Zool. J. Linn. Soc. 2009, 157, 51–69. [Google Scholar] [CrossRef] [Green Version]
- Deichmann, E. The Holothurioidea collected by the Velero III and IV during the years 1932 to 1954. Part II. Aspidochirota. Allan Hancock Pac. Exped. 1958, 11, 253–349. [Google Scholar]
- Michonneau, F.; McPherson, S.; O’Loughlin, P.M.; Paulay, G. More than meets the eye: Diversity and geographic patterns in sea cucumbers. arXiv 2015, arXiv:014282. [Google Scholar] [CrossRef] [Green Version]
- Michonneau, F. Cryptic and not-so-cryptic species in the complex “Holothuria (Thymiosycia) impatiens” (Forsskål, 1775) (Echinodermata: Holothuroidea: Holothuriidae). arXiv 2015, arXiv:014225. [Google Scholar] [CrossRef] [Green Version]
- WoRMS—World Register of Marine Species. Available online: https://www.marinespecies.org/aphia.php (accessed on 4 February 2023).
- Dabbagh, A.R.; Keshavarz, M. Three sea cucumbers from the Bandar-E Bostaneh coast (Persian Gulf, Iran). World Appl. Sci. J. 2011, 13, 1933–1937. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Rasolofonirina, R.; Jangoux, M. Appearance and development of skeletal structures in Holothuria scabra larvae and epibiont juveniles. SPC Beche-de-Mer Inf. Bull. 2005, 22, 5–10. [Google Scholar]
- Hoareau, T.B.; Boissin, E. Design of phylum-specific hybrid primers for DNA barcoding: Addressing the need for efficient COI amplification in the Echinodermata. Mol. Ecol. Resour. 2010, 10, 960–967. [Google Scholar] [CrossRef]
- Arndt, A.; Marquez, C.; Lambert, P.; Smith, M.J. Molecular Phylogeny of Eastern Pacific Sea Cucumbers (Echinodermata: Holothuroidea) Based on Mitochondrial DNA Sequence. Mol. Phylogenet. Evol. 1996, 6, 425–437. [Google Scholar] [CrossRef]
- Kerfeld, C.A.; Scott, K.M. Using BLAST to Teach “E-value-tionary” Concepts. PLoS Biol. 2011, 9, e1001014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Wangüemert, M.; Domínguez-Godino, J.A.; Cánovas, F. New records of sea cucumbers inhabiting Mar Menor coastal lagoon (SE Spain). Mar. Biodiv. 2018, 48, 2177–2182. [Google Scholar] [CrossRef]
- So, J.J.; Mercier, A.; Hamel, J.-F. Habitat utilisation, growth and predation of Cucumaria frondosa: Implications for an emerging sea cucumber fishery. Fish. Manag. Ecol. 2010, 17, 473–484. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. popart: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Nei, M. Molecular Evolutionary Genetics; Columbia University Press: New York, NY, USA, 1987; p. 512. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. jModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- James, D.B. Taxonomic Studies of the Species of Holothuria (LINNAEUS, 1767) from the Seas Around India 505 1 Part I. 1992. Available online: http://eprints.cmfri.org.in/7434/1/449J._BOMBAY_NAT.HISTORY_SOC_1995.pdf (accessed on 1 January 2020).
- Raymund, D.J.; Victoria, T.N.; Vinya, M.A.; Frederick, G.T. The Body Wall Spicule Formations of Mature Holothuria impatiens Found in La Union, Philippines. J. Nat. Allied Sci. 2019, 3, 42–45. [Google Scholar]
- Ramofafia, C.; Battaglene, S.C.; Bell, J.D.; Byrne, M. Reproductive biology of the commercial sea cucumber Holothuria fuscogilva in the Solomon Islands. Mar. Biol. 2000, 136, 1045–1056. [Google Scholar] [CrossRef]
- Mercier, A.; Sun, Z.; Baillon, S.; Hamel, J.-F. Lunar rhythms in the deep sea: Evidence from the reproductive periodicity of several marine invertebrates. J. Biol. Rhythm. 2011, 26, 82–86. [Google Scholar] [CrossRef]
- Drumm, D.J.; Loneragan, N.R. Reproductive biology of Holothuria leucospilota in the Cook Islands and the implications of traditional fishing of gonads on the population. J. Mar. Freshw. Res. 2005, 39, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Dabbagh, A.-R.; Keshavarz, M.; Mohammadikia, D.; Afkhami, M.; Nateghi, S.A. Holothuria scabra (Holothuroidea: Aspidochirotida): First record of a highly valued sea cucumber, in the Persian Gulf, Iran. Mar. Biodivers. Rec. 2012, 5, e69. [Google Scholar] [CrossRef]
- Wesselmann, M.; Gonzalez-Wanguemert, M.; Serrao, E.A.; Engelen, A.H.; Renault, L.; Garcia-March, J.R.; Hendriks, I.E. Genetic and oceanographic tools reveal high population connectivity and diversity in the endangered pen shell Pinna nobilis. Sci. Rep. 2018, 8, 4770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, L.; Adcock, G.J.; Smith, P.J.; Ramírez, J.H.B.; Carvalho, G.R. Loss of Microsatellite Diversity and Low Effective Population Size in an Overexploited Population of New Zealand Snapper (Pagrus auratus) | PNAS. Available online: https://www.pnas.org/doi/full/10.1073/pnas.172242899 (accessed on 4 February 2023).
- Hamamoto, K.; Soliman, T.; Poliseno, A.; Fernandez-Silva, I.; Reimer, J.D. Higher Genetic Diversity of the Common Sea Cucumber Holothuria (Halodeima) atra in Marine Protected Areas of the Central and Southern Ryukyu Islands. Front. Conserv. Sci. 2021, 2, 736633. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Body Size | Rg1 | Rg2 | Rg3 |
|---|---|---|---|
| Mid-dorsal (µm) | |||
| Buttons’ length | 69.5–97.2 | 63.8–77.4 | 65.7–80.4 |
| Buttons’ width | 31–32.3 | 31.6–35 | 29.5–34.9 |
| Tables’ disc diameter | 78.1–83.6 | 84.7–91.9 | 74.7–95.8 |
| Mid-ventral (µm) | |||
| Buttons’ length | 70.9–73.7 | 73–77.9 | 67.9–78.1 |
| Buttons’ width | 30.8–32.4 | 28.5–36.6 | 32.3–33 |
| Tables’ disc diameter | 85.9–99 | 84.7–85.3 | 81.8–89.2 |
| Species | GenBank Code | Location/Voucher | References |
|---|---|---|---|
| Holothuria (Thymiosycia) impatiens | GQ214760 | Ho273/Cabo de Palos, Spain | [2] |
| Holothuria impatiens | KX384727 | Mar Menor, Spain | [27] |
| Holothuria (Thymiosycia) impatiens | GHI07432 | Gulf of Tunis, S1 GHI1 | Current study |
| GHI09431 | Gulf of Tunis, S1 GHI2 | ||
| GHI10433 | Gulf of Tunis, S1 GHI3 | ||
| GHI11434 | Gulf of Tunis, S1 GHI4 | ||
| GHI12435 | Gulf of Tunis, S1 HIT1 | ||
| GHI13436 | Gulf of Tunis, S1 HIT2 | ||
| GHI14437 | Gulf of Tunis, S1 HIT3 | ||
| GHI15438 | Gulf of Tunis, S1 HIT4 | ||
| GHI16439 | Gulf of Tunis, S1 HIT5 | ||
| GHI18430 | Gulf of Tunis, S1 GHI5 | ||
| GHI01431 | Gulf of Tunis, S1 GHI6 | ||
| GHI02432 | Gulf of Tunis, S2 HITT1 | ||
| GHI03433 | Gulf of Tunis, S2 HITT2 | ||
| GHI04434 | Gulf of Tunis, S2 HITT3 | ||
| GHI05435 | Gulf of Tunis, S2 HITT4 | ||
| GHI06436 | Gulf of Tunis, S2 HITT5 | ||
| GHI17439 | Gulf of Tunis, S2 HITT6 | ||
| Holothuria (Thymiosycia) impatiens | MOLAF0087 MOLAF124 MOLAF0123 | Italy | [19] |
| Holothuria impatiens | GQ920687 GQ920688 | Golfed’Aqaba, Red Sea, Egypt | - |
| Cucumaria frondosa | EU811805 | LB2CH22/Canada | [28] |
| Samples | Weight, W (g) | Length, L (cm) |
|---|---|---|
| GHI05435 * | 26.14 | 11 |
| GHI06436 | 2.93 | 5 |
| GHI03433 * | 16.58 | 9.5 |
| GHI02432 * | 7.18 | 6 |
| GHI1431 * | 5.34 | 6.5 |
| GHI18430 * | 5.27 | 7 |
| GHI15438 * | 7.65 | 6.3 |
| GHI14437 | 3.87 | 5.5 |
| GHI12435 | 0.92 | 2.3 |
| GHI11434 | 1.16 | 2.1 |
| GHI10433 * | 15.7 | 8.6 |
| GHI09431 | 3.02 | 4.7 |
| GHI07432 * | 27.09 | 10 |
| GHI13436 * | 3.55 | 3 |
| GHI16439 * | 9.85 | 7.5 |
| GHI17439 * | 11.69 | 8 |
| GHI04434 | 3 | 2.5 |
| Red Sea | Mediterranean | Tunisia | Italy | Spain | |
|---|---|---|---|---|---|
| Red Sea | 0.0000 | ||||
| Mediterranean | 0.2019 | 0.0032 | |||
| Tunisia | 0.2021 | na | 0.0015 | ||
| Italy | 0.2026 | na | 0.0022 | 0.0039 | |
| Spain | 0.2039 | na | 0.0032 | −0.0005 | 0.0059 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalthoumi, D.; Francisco, S.M.; Miladi, M.; Ruiz-Canales, A.; Azzouna, A.; Robalo, J.I. The First Report of Holothuria (Thymiosycia) impatiens (Forsskål, 1775), (Holothuroidea: Holothuriidae) from Tunisia (Mediterranean Sea): Taxonomic, Morphological, and Molecular Data Compilation. Diversity 2023, 15, 542. https://doi.org/10.3390/d15040542
Kalthoumi D, Francisco SM, Miladi M, Ruiz-Canales A, Azzouna A, Robalo JI. The First Report of Holothuria (Thymiosycia) impatiens (Forsskål, 1775), (Holothuroidea: Holothuriidae) from Tunisia (Mediterranean Sea): Taxonomic, Morphological, and Molecular Data Compilation. Diversity. 2023; 15(4):542. https://doi.org/10.3390/d15040542
Chicago/Turabian StyleKalthoumi, Donia, Sara M. Francisco, Malek Miladi, Antonio Ruiz-Canales, Atf Azzouna, and Joana I. Robalo. 2023. "The First Report of Holothuria (Thymiosycia) impatiens (Forsskål, 1775), (Holothuroidea: Holothuriidae) from Tunisia (Mediterranean Sea): Taxonomic, Morphological, and Molecular Data Compilation" Diversity 15, no. 4: 542. https://doi.org/10.3390/d15040542