The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta



1
Taxonomy and Systematics Group, Naturalis Biodiversity Center, P.O. Box 9517, 2300 Ra Leiden, The Netherlands
2
Research Center for Oceanography, Indonesian Institute of Sciences (LIPI), Jl. Pasir Putih I, Ancol Timur, Jakarta 14430, Indonesia
*
Author to whom correspondence should be addressed.
Diversity 2019, 11(3), 46; https://doi.org/10.3390/d11030046
Submission received: 30 January 2019
/
Revised: 16 March 2019
/
Accepted: 18 March 2019
/
Published: 21 March 2019
(This article belongs to the Special Issue Cross-shelf Variation in the Structure and Function of Coral Reef Assemblages)
Abstract
:Many coral reef systems are shelf-based and consist of reefs that are arranged in rows parallel to the coastline. They usually show an increase in species richness in the offshore direction, coinciding with decreasing terrigenous impact and a deeper seafloor. These two conditions usually concur, which makes it less easy to distinguish how each of them influences coral diversity separately. Since reefs off Jakarta (in the Thousand Islands archipelago) are arranged in an 80 km long string perpendicular to the coastline in south-to-north direction, with a maximum shelf depth halfway along (instead of at the end of) the string, this archipelago is very suitable for studies on inshore–offshore gradients. In the present study, mushroom corals (Fungiidae; n = 31) were used to examine diversity patterns on 38 reef sites along such a gradient, involving species richness over their entire depth range from reef flat to reef base (2–30 m) and separately at shallow depths (2–6 m). Total species diversity was highest in the central part of the archipelago, with unique species occurring in deep habitats. Diversity at shallow depths was only slightly higher here than at reefs located more nearshore and offshore, which both had less clear water. Therefore, shelf depth and distance from the mainland can be considered separate determinants of coral diversity off Jakarta.
Keywords:
Scleractinia; Fungiidae; Indonesia; Java Sea; mega city; latitude; river outlets; water transparency; blast fishing1. Introduction
Shelf-based reef systems are ideal model areas in which to study the influence of inshore-to-offshore environmental gradients on the diversity of reef coral assemblages. Reefs located closer to major shorelines (hereafter inshore or nearshore) are exposed to terrigenous and anthropogenic impact from nearby land mass [1] and to oceanic conditions at the offshore side, resulting in increasing water quality away from the coastline [2,3,4,5,6,7]. In addition, the seafloor of such shelf systems usually shows a gradually increasing depth in the same direction, up to the shelf ridge [8,9]. Nearshore reef zones receive sediment from river outlets, which keeps the seafloor shallow and water more turbid here [10]. In order to examine the effects of water quality and depth on species diversity, they should be analyzed separately. To serve as a model reef system and to enable the distinction of clear diversity patterns, the reefs should be abundant, evenly distributed, and be geomorphologically uniform. Most such reef systems are predominantly arranged alongside and parallel to the mainland shoreline, such as the Great Barrier Reef, located off the north-east Australian coast [11,12,13,14,15], and various smaller shelf-based reef complexes, such as the Spermonde Archipelago off Southwest Sulawesi [5,16,17,18,19], the Padang shelf reef system off West Sumatra [20,21], the Berau Archipelago off eastern Borneo [22,23,24], the Semporna barrier reef at Northeast Borneo [25], the patch reefs off Brunei and Sarawak at Northwest Borneo [26,27,28], the barrier reef system along the coast of Belize [29,30], Madang Lagoon in northern Papua New Guinea [31,32], the southwestern lagoon of New Caledonia [33,34], and a few reef complexes along the Saudi Arabian coastline of the Red Sea [35,36,37]. Many of these examples are sheltered from deeper water (>200 m depth) by a chain of shallow barrier reefs, with the exception of West Sumatra, where reefs are separated from the ocean by a series of large islands [20,21].
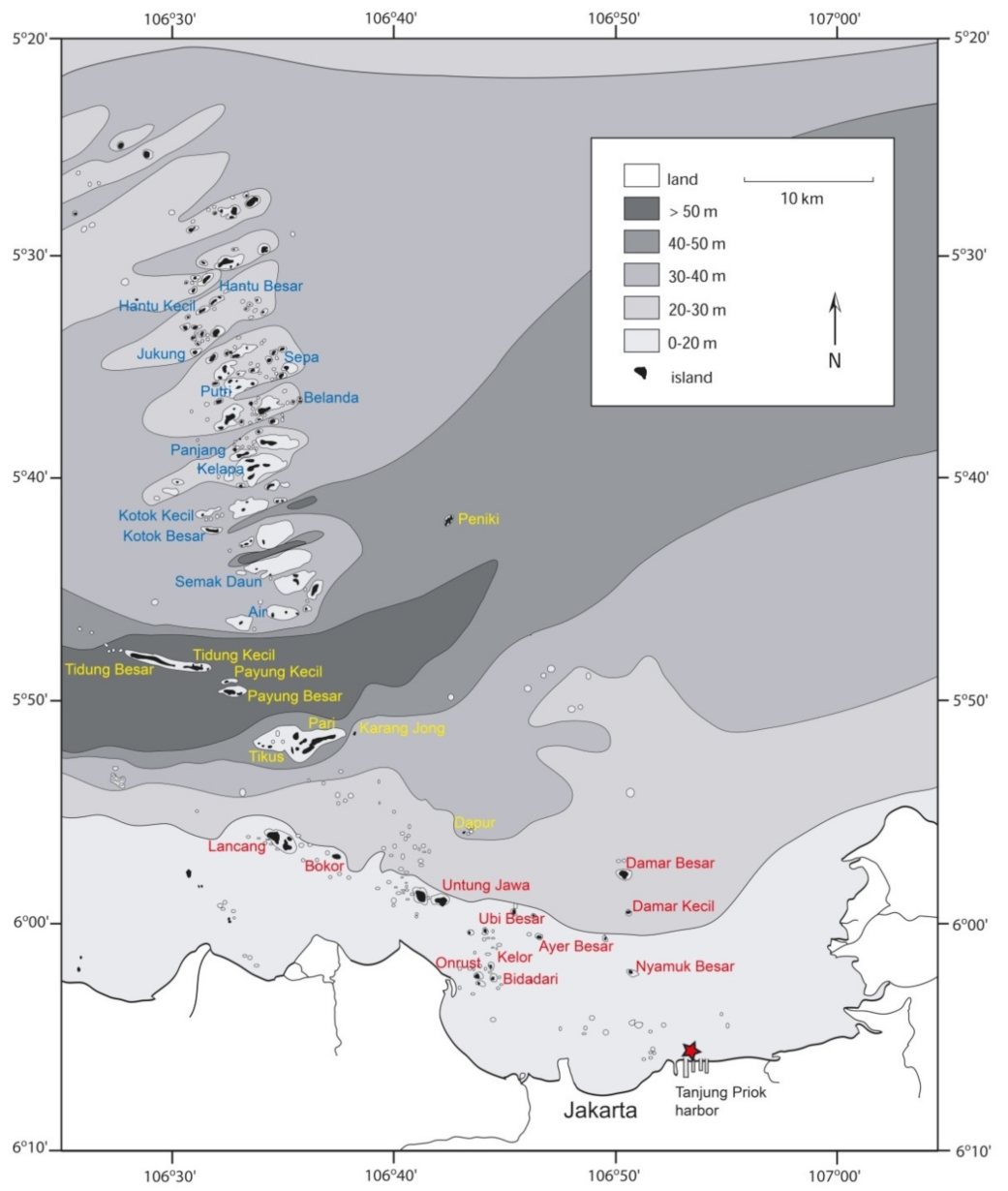
In contrast, the Thousand Islands off Jakarta (locally known as Kepulauan Seribu) is a string of patch reefs (80 km long) oriented perpendicularly to the coastline from south to north in the shallow Java Sea, which is surrounded by the land masses of Java, Sumatra, and Borneo. The shelf is deepest in the middle of the archipelago (Figure 1). A river used to run here during Pleistocene low sea level stands, which now acts as a sunken channel with a predominantly westward current [38,39,40,41,42]. The islands are cay-crowned patch reefs [43,44], most of which are inhabited by fishermen or used for recreational purposes [45,46,47,48,49]. Some cays in Jakarta Bay have disappeared due to dredging and subsidence caused by groundwater extraction [50,51,52,53]. A large part of the bay within the present 10 m isobath will become reclaimed land, causing a few reefs to disappear and others to become situated closer to the coastline [54,55]. Consequently, nearshore reefs are exposed to sedimentation as a result of dredging activities, which is harmful to most corals [56]. Owing to the proximity of Jakarta, a large conurbation and a major port, reef communities here are also impacted by various kinds of pollution [1,57,58,59,60], much of which comes from river outlets inside and close to Jakarta Bay [51,61,62]. The reef communities here are threatened, because anthropogenic stresses have already been blamed for the disappearance of reef-dwelling species in Jakarta Bay during the last century [63,64]. On the positive side, the reefs off Jakarta have recovered from coral bleaching [65,66] and they survived attacks by the corallivorous crown-of-thorns sea star [67,68,69].
Since shelf depth in the Thousand Islands reef complex is largely independent of distance offshore, unlike the shelf-based reef complexes mentioned earlier, this archipelago is suitable for studies on the effect of seafloor depth on the diversity of reef-dwelling species. Reefs based on a deep seafloor have deep-ranging reef slopes and reef bases, offering habitats to many species that are absent in shallower water. Species dwelling on deeper reef parts receive less light and are less exposed to wave action than species dwelling on reef flats and upper reef slopes [70,71,72]. Thus, these reefs are expected to have a relatively rich coral fauna by also offering habitats to species that prefer deep reef conditions.
The Indo-Pacific mushroom coral family Fungiidae (Scleractinia) is an ideal model group in which to study cross-shelf diversity patterns because they are abundant from nearshore to offshore and from shallow to deep habitats [17,73,74], which include mesophotic depths [75,76,77]. Most species (80%) are free-living, which enables them to colonize unconsolidated substrates, such as sandy bottoms and rubble [71,78,79,80]. With over 50 valid species, this family forms a large taxon, representing about 8% of all Indo-Pacific reef-inhabiting scleractinians [81]. In addition to free-living mushroom corals, there are foliaceous species and encrusting ones [82,83]. Over 60% of all species represent solitary forms with a single mouth, most of which remain small, whereas the largest species can have more than one mouth [84,85,86]. During earlier studies in the Bay of Jakarta/Thousand Islands, 24 fungiid species were found at depths of 21 m or less [66,87,88,89,90]. Fungiidae do not show notably abnormal patterns in beta-diversity studies compared to other scleractinian families [3,88,91,92]. In biodiversity surveys off Borneo, Fungiidae showed similar species richness accumulation curves as the families Agariciidae and Euphylliidae [25,93,94]. Although the family Fungiidae has several species with the ability to live at depths >30 m [17,74,75,76,77], the same goes for staghorn corals of the family Acroporidae [94,95,96,97].
In the present study, Fungiidae are utilized as a model taxon to examine how coral diversity at shallow depths (2–6 m on reef flats and upper slopes) and over the whole reef profile (2–30 m from the reef flat down to the reef base) varies over an inshore–offshore gradient and over increasing depth on the seafloor off Jakarta. Previous coral diversity surveys in the same research area (in 1985 and 1995) were limited to shallow depths only (1–5 m) and thus excluded coral species possibly occurring at greater depths [88,89]. Distance from the mainland represents reach of terrigenous impact, which is most clearly visible through the presence of river plumes [24,64]. Shelf depth can influence wave-driven sediment resuspension [98,99], with relatively clear water expected above deep seafloors, and also on the maximum depth range of coral assemblages, including species that are specialized in deep-water habitats. A diversity gradient at shallow depths alone may also depend on abiotic parameters related to distance from the shoreline and to maximum shelf depth, for example by varying species concentrations that are partly determined by the distribution of rare species. The reef complex off Jakarta, with a shelf depth not linearly related to distance offshore, is therefore suitable for testing whether these two parameters can operate as distinct drivers of coral diversity.
2. Materials and Methods
2.1. Research Area and Abiotic Parameters
Field data were sampled in September 2005 during a survey in Jakarta Bay and the Thousand Islands, together forming an 80 km long string of islands more or less perpendicular to the coastline in NNW direction (Figure 1). The expedition was organized by the Research Centre for Oceanography (Indonesian Institute of Sciences = LIPI) in Jakarta and by Naturalis Biodiversity Center, Leiden [100]. Thirty-eight reef sites were visited, mostly located at the northwestern side of each island, similar to expeditions in 1985 and 1995 [2,101,102,103] but now including three extra reefs: Karang Jong, Pulau Peniki, and an unnamed shoal south of Pulau Pari (Figure 1, Table 1).
The 38 reef sites were divided over three zones arranged from inshore to offshore, varying in shelf depth: 11 sites in Zone 1 (0–30 m), 15 sites in Zone 2 (30–60 m), 12 sites in Zone 3 (20–40 m). Seafloor depth was determined with the help of nautical charts (scale 1:20,000 or 1:50,000) issued by the Indonesian Navy [104,105,106,107,108,109,110,111]. Transparency of the water column was measured with a standard 30 cm wide secchi disc with maximum visibility depth marked in meters along a rope [111]. Site coordinates were determined with a GPS (Garmin eTrex). Coordinates of reference points on the coastline, representing possible sources of pollution (mainland, river outlets, and Tanjung Priok harbor), were found with Google Maps [112]. To convert coordinate units from DMS (degrees, minutes, seconds) to DD (decimal degrees) and to measure the distance from each reef site to reference points on the mainland, the program GPS Coordinates was used [113].
2.2. Coral Data
At each site a distinction was made between mushroom coral species recorded at 2–6 m depth only (reef flat and upper slope) and those over the whole reef profile, from 2 m depth down to the reef base at 30 m depth or less. These separate approaches (shallow vs. all depths) allow a distinction between coral diversity depending on water quality (distance offshore, turbidity) and diversity depending on bathymetrical reef range (limited by maximum shelf depth). The coral data from shallow depths were obtained from 30 m long, 1 m wide belt transects, three at 3 m and three at 5 m [100,114], within a depth range of 2–6 m, while records from deeper water were acquired by the roving diver technique [115]. Each data sample (one per reef site) is based on a single 60 min dive.
Representative specimens were photographed with a Ricoh Caplio RR30 camera in a Sea&Sea DX-3000 housing. Identifications were based on taxonomic revisions and faunistic works available in 2005 [90,116]; their present classifications follow subsequent species descriptions and taxonomic revisions [82,83,85,86,117]. Earlier records [66,90] and voucher specimens in the coral collection of Natural Biodiversity Center from the research area were reexamined to check for species that could have been missed during the 2005 survey.
2.3. Data Analysis
Abiotic parameters and diversity counts were correlated by linear and non-linear regressions and fitted line plots through quadratic and linear model functions in the Minitab package [118]. A quadratic model was preferred above a linear model when it resulted in a better correlation (higher r-value). The following abiotic relations were analyzed: Shelf depth–latitude, secchi depth–distance to mainland, and secchi depth–shelf depth. Total mushroom coral diversity was correlated with shelf depth, secchi depth, latitude, distance to mainland, distance to harbor, and distance to nearest river outlet. Because the last four functions were very similar in correlation, diversity in shallow water was only related to shelf depth, secchi depth, and latitude.
Species richness estimators were used to compare the total number of observed species (incidence data) for all 38 sites together with theoretically expected species numbers in order to determine whether the diversity data set is representative. The analyses were performed with the program EstimateS 9.1 [119]. The observed number of species (SObs) is presented as an asymptotic accumulation curve, which was constructed after the sample order was randomized and the values were averaged, resulting in a mean and standard deviation of the observed species number for each consecutive sample unit and for the estimators Chao2 and ICE (incidence coverage-based estimator), the latter of which is more sensitive to unique species. Therefore the number of uniques (species represented in a single sample) is also given.
3. Results
3.1. Abiotic Parameters
Shelf depth does not show a linear correlation with latitude (r = 0.385, p > 0.05). A deep channel (Zone 2) roughly runs in east-to-west direction and separates the two shallow shelf zones, Zone 1 and Zone 3 (Figure 1). This bathymetric variation is also reflected in the maximum shelf depth for each site, with 54 m depth records at Payung Besar and Payung Kecil (Figure 2a; Table 1).
Secchi depth shows a similar relation, with the clearest water in the mid-section of the study area (Zone 2), where the seafloor is deepest (Figure 1b and Figure 2b; Table 1). Therefore a quadratic model shows a stronger correlation between secchi depth and distance to mainland (r = 0.822) than a linear model (r = 0.561), meaning that water is less transparent north and south of the channel. Secchi depth shows a very significant linear relation with shelf depth (p < 0.00001), which is consistent with the clearest water occurring near reefs of Zone 2 (Figure 2c).
3.2. Species Diversity Patterns
3.2.1. Entire Depth Range
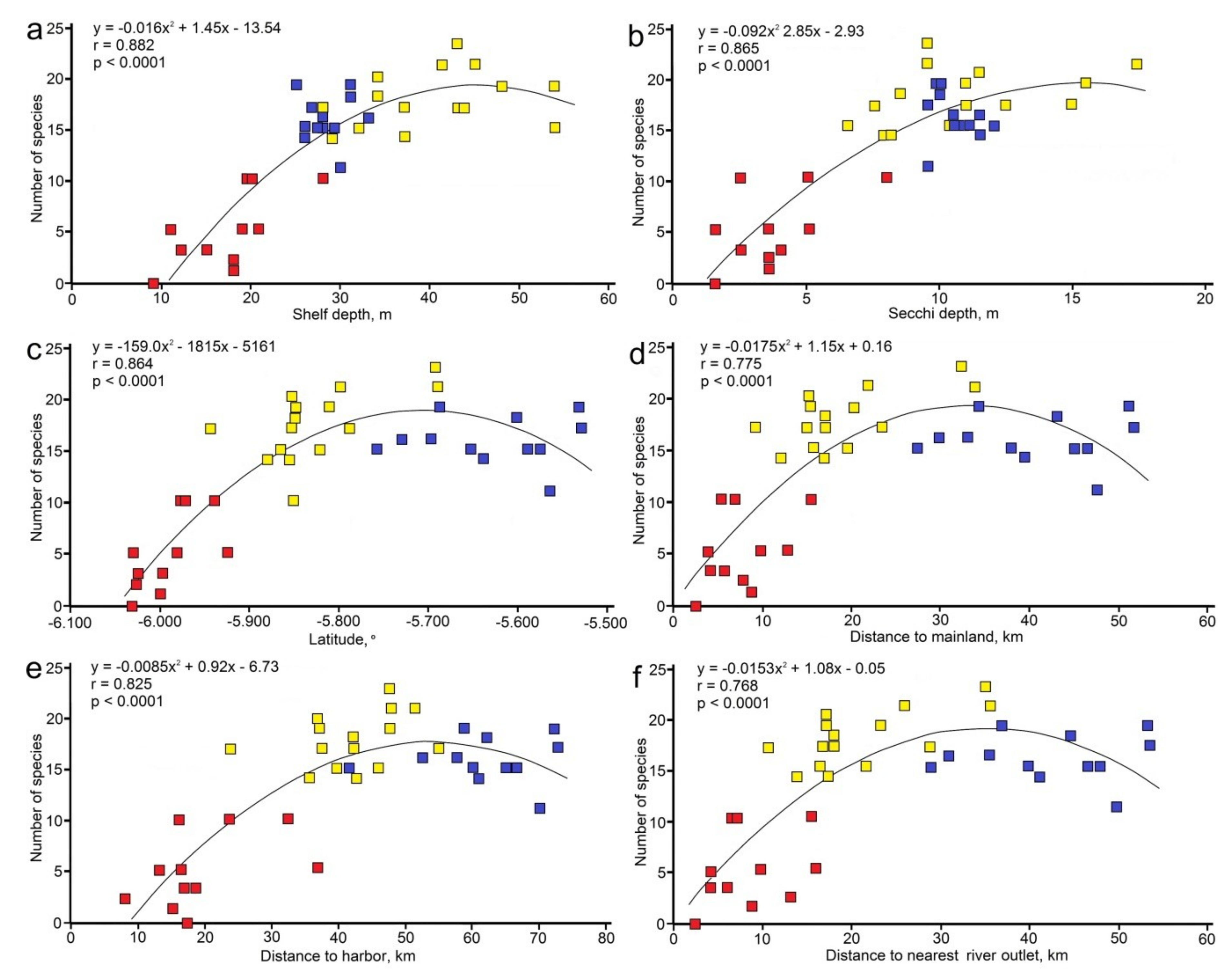
The number of mushroom coral species per site varied from 0 at Pulau Onrust (Bay of Jakarta) to 23 at Pulau Peniki in Zone 2 (Table S1; Figure 3a). The significant relation between species number and shelf depth fits better in a quadratic model (r = 0.882) than in a linear model (r = 0.772).
The function approaches an asymptote, suggesting a saturation effect, meaning that over a shelf depth of ca. 42 m no higher numbers of observed species per sample unit can be expected. The number of species also depends on water transparency (secchi depth, Figure 3b), with a slightly stronger correlation for the quadratic model (r = 0.865) than the linear model (0.820) and a saturation point at ca. 16 m visibility depth. The quadratic relations between species diversity and distances of the reef sites from the mainland (r = 0.775), nearest river outlet (r = 0.768), and Tanjung Priok harbor (r = 0.825) are very similar to such a relation with latitude (r = 0.864), with the last being the strongest (Figure 3c–f). Their r-values do not differ significantly after Fisher r-to-z transformation (p > 0.05). All fit better than linear relations (r = 0.559, 0.574, 0.697, 0.655, respectively) and reflect a maximum species diversity in Zone 2.
A larger number of species in Zone 2 is also shown by the total number of all observed species in that zone (n = 29), as compared to nearshore Zone 1 (n = 16) and offshore Zone 3 (n = 24). With the addition of two historical records, the total numbers for Zone 2 and Zone 3 are each one species higher (Table 2; Figure 4d–f). Seven species represent new records for the research area (Table 2).
Some species are common in all shelf zones, such as the free-living Fungia fungites, Pleuractis paumotensis, and Sandalolitha robusta, and the attached species Lithophyllon undulatum (Table 2). A few species with shallow-water records have only been encountered rarely in Zone 2 and Zone 3, such as the free-living species Halomitra pileus, Lithophyllon scabra, Lobactis scutaria, and Pleuractis granulosa, and the attached Podabacia kunzmanni (Table 2; Figure 4). Typical taxa for these two zones are the free-living Ctenactis albitentaculata, Ctenactis crassa, Cycloseris boschmai, Cycloseris costulata, Cycloseris cyclolites, and Sandalolitha dentata (Table 2; Figure 5). Deep-living species were found on lower reef slopes, such as the encrusting species Cycloseris explanulata and Cycloseris mokai, and on reef bases in both zones, such as the free-living Cycloseris fragilis, Cycloseris sinensis, and Cycloseris vaughani (Table 2; Figure 6). Cycloseris fragilis and C. sinensis were found in complete shape and in self-fragmenting mode (Figure 6c,d).
3.2.2. Shallow Depths
Species richness at 2–6 m depth excludes species only found on lower reef slopes and sandy reef bases. The relation between species number and shelf depth is represented better by a quadratic model (Figure 7a; r = 0.795) than a linear model (r = 0.614). The maximum values are found in Zone 3, while the plot reaches its maximum in Zone 2. A similar relation is found between species richness at 2–6 m depth and secchi depth, showing an increasing species richness with higher visibility from Zone 1 to Zone 2 (Figure 7b; quadratic r = 0.829, linear r = 0.735). The number of shallow species also increases in northward direction with distance offshore, approaching an asymptotic relation (Figure 7c; quadratic r = 0.852, linear r = 0.760). Totals of shallow-water records among the three zones vary little with values of 16, 18, and 17, respectively (Table 2).
At some offshore localities, such as Pulau Putri in Zone 3, reefs were damaged at shallow depths because of blast fishing. Living mushroom corals were found in between dead and live coral fragments of other species (Figure 8).
3.2.3. Species Richness Estimators
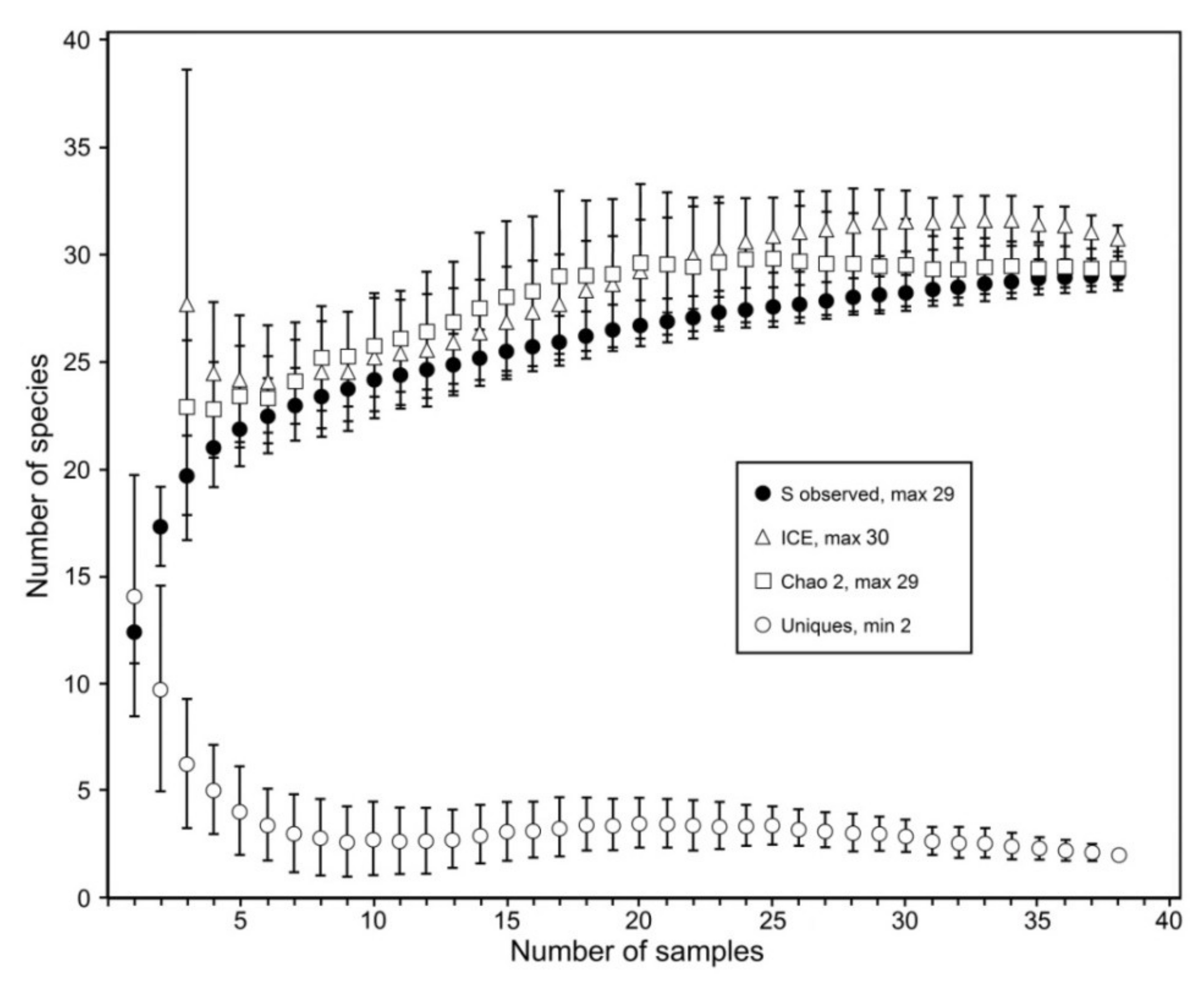
Application of species richness estimators (all reaching asymptotes) indicates that the total observed number of species (n = 29) is very similar to the estimated numbers Chao2 (n = 29) and ICE (n = 30) (Figure 9). The number of uniques (n = 2) is low. Two additional species observed in 1983 and 1995, respectively, were not taken into account.
4. Discussion
The arrangement of the reefs off Jakarta in an 80 km long string perpendicular to the coastline makes this archipelago suitable for a study of inshore–offshore diversity patterns. The present results show that the highest diversity of mushroom corals off Jakarta can be found on the reefs surrounded by a deep seafloor in the central section of the archipelago and not on the northernmost reefs, which are farthest away from the mainland. This concerns diversity at shallow depths, probably due to clear water in the central part, and over the entire reef profile, including a sandy base with sediment-tolerant species. The offshore reefs have shown a large reduction in water transparency since the 1920s, while the turbidity on the inshore reefs has not changed much until 2005, after which offshore reefs started to show a better visibility compared to inshore reefs [57,120,121,122]. Since the offshore reefs of Zone 3 are based on a shallow seafloor, sediment from the bottom may become resuspended more easily by wave action here than near reefs surrounded by deep water (Zone 2).
Lobactis scutaria, uniquely found in Zone 2 (Table 2; Figure 7b), is an example of a species that is common in clear water at shallow depths [16]. Other fungiids exclusively occurring in Zone 2 are free-living species found on reef slopes, such as C. boschmai and C. cyclolites (Figure 8c,d), and a single species living on a deep reef base, C. vaughani (Figure 6f), which is consistent with their distribution patterns at SW Sulawesi [17]. The latter species is also known to occur at mesophotic depths [77]. Encrusting species usually found on lower reef slopes, such as C. explanulata and C. mokai, were also most frequently observed in Zone 2. At SW Sulawesi, the outermost reefs are situated on a shallow barrier, which are less rich in fungiids than mid-shelf reefs surrounded by deeper reef slopes and reef bases [1,17,74], which is consistent with the depth-related diversity off Jakarta. Interestingly, mushroom corals in zones 2 and 3 may not be abundant at very shallow depths (<2 m), where families like Acroporidae and Poritidae are dominant [123].
Most previous coral studies on inshore–offshore gradients near Jakarta did not focus on diversity but on coral cover at shallow depths, generally finding highest values in Zone 2 and Zone 3 [2,57,58,114,124]. Reports dealing with coral diversity were limited to observations at shallow depths (5 m), and also with high numbers in Zone 2 and Zone 3 [88,89,102,125], thus discounting species that prefer greater depths. In a parallel study (in 2005) on algae down to 30 m depth, a maximum diversity was also found in the deeper part of the shelf (with the addition of some reefs more to the north) and a declining species number towards the north [126]. On the other hand, fish counts in shallow water (<10 m depth) resulted in an increase of species richness with distance offshore [127]. A similar relation was found for sponges collected from depths <20 m [128]. However, these studies did not distinguish between taxa that prefer shallow habitats and those that usually live deeper.
The present study did not reveal significant differences among species diversity gradients along a latitudinal gradient, distance offshore, distance from the nearest river outlet, and distance from Tanjung Priok harbor (Figure 2). There are multiple river outlets distributed along the coastline inside and outside Jakarta Bay, which together cause a diffuse influx of freshwater and contaminants along the shoreline [122,129], whereas reefs outside the Bay of Jakarta, in particular those farthest away from the mainland, show little difference in terrigenous impact [58].
A marine park has been designed to include only some of the most offshore reefs [45,47,130,131]. These remote reefs needed protection because illegal blast fishing was a very common practice here until the mid-1980s [132,133]. Its impact could still be witnessed in 2005 (Figure 8) and a recent study indicated that, until recently, blast fishing has still been going on [49]. Blast-generated craters and rubble fields may take decades to recover [133,134,135,136] and while it appears that free-living mushroom corals among the dead coral fragments have survived, they may also have arrived by migration [137]. Although the blast fishing predominantly took place at shallow depths, there are no signs that this affected species richness here in 2005.
The 2005 survey resulted in seven new records of reef coral species for Jakarta, including one from 4 m depth, the small attached Podabacia kunzmanni, which was described in 2009 and previously not recognized [82], and two small free-living species from deep (>20 m) sandy reef bases, C. sinensis and C. vaughani (Table 2). With a record of 31 fungiids, the islands of Jakarta are not as diverse as most areas studied in the adjacent Coral Triangle (30 ≤ n ≤ 44) [25,42,75,138] but they are more diverse than reef areas in the nearby Gulf of Thailand (20 ≤ n ≤ 28) [139].
Several corals of C. fragilis and C. sinensis were showing autotomy (self-fragmentation) and regeneration on the deep sandy reef bases as a mechanism of asexual reproduction (Figure 6c,e). This has not reported before from the Thousand Islands but it is common in similar habitats elsewhere [77,140,141]. On sandy substrates, this mechanism may replace sexual reproduction, for which a solid settlement substrate is needed before the corals become free-living [142,143]. Small, unattached mushroom corals can survive on sandy substrates because they are able to shed sediments [144,145] and they can move themselves away from direct threats [146,147,148]. Nevertheless, unattached fungiids are not unique as free-living and mobile corals on reefs worldwide and in the fossil record [78,79,80,149,150], which implies that the Fungiidae still constitute a suitable model taxon for Indo-Pacific coral diversity surveys.
Since many studies on reef condition focus on shallow reef zones, the present results indicate that more attention should be given to deeper reef parts. Recent research on reefs and reef zones > 20 m in depth has resulted in additional species records for other reef coral faunas, such as in the Persian Gulf [141], Taiwan [151], and the Great Barrier Reef [95]. Biodiversity surveys should therefore not be limited to shallow depths. There may be many other influences on species diversity, which have not been considered in the present study, such as salinity [24], dominant substrate type [17], and exposure to wave action and currents [1,17,91]. Future studies in the research area might focus on the impact of these natural factors on coral diversity and also on the effect of anthropogenic impact, such as land reclamation, pollutants, and worsening water quality [1,5,7].
Supplementary Materials
The following is available online at https://www.mdpi.com/1424-2818/11/3/46/s1, Table S1: Jakarta mushroom coral species records.
Author Contributions
Conceptualization, B.W.H., G. and S.; methodology, B.W.H., G. and S.; validation, B.W.H. and G.; formal analysis, B.W.H. and G.; investigation, B.W.H. and G.; resources, B.W.H. and S.; data curation, B.W.H. and G.; writing—original draft preparation, B.W.H.; writing—review and editing, B.W.H.; visualization, B.W.H.; supervision, B.W.H. and S.; project administration, B.W.H. and S.; funding acquisition, B.W.H. and S.
Funding
This research was funded by the Council for Earth and Life Sciences of the Netherlands Organisation for Scientific Research (ALW-NWO grant 852.000.50).
Acknowledgments
The authors thank Yosephine Tuti and other expedition participants (RCO-LIPI and Naturalis) for logistic support. The first author is grateful to the Indonesian Institute of Science (LIPI, Jakarta) for the research permit. Erik-Jan Bosch (Naturalis) made the original version Figure 1. We thank two anonymous reviewers for their constructive comments and the editors for inviting us to participate in the special issue “Cross-shelf Variation in the Structure and Function of Coral Reef Assemblages”.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Heery, E.C.; Hoeksema, B.W.; Browne, N.K.; Reimer, J.D.; Ang, P.O.; Huang, D.; Friess, D.A.; Chou, L.M.; Loke, L.H.L.; Saksena-Taylor, P.; et al. Urban coral reefs: Degradation and resilience of hard coral assemblages in coastal cities of East and Southeast Asia. Mar. Pollut. Bull. 2018, 135, 654–681. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; Becking, L.E.; de Voogd, N.J.; Renema, W.; de Beer, M.; van Soest, R.W.M.; Hoeksema, B.W. Variation in the diversity and composition of benthic taxa as a function of distance offshore, depth and exposure in the Spermonde Archipelago, Indonesia. Estuar. Coast. Shelf Sci. 2005, 65, 557–570. [Google Scholar] [CrossRef]
- Cleary, D.F.R.; DeVantier, L.; Vail, L.; Manto, P.; de Voogd, N.J.; Rachello-Dolmen, P.G.; Tuti, Y.; Budiyanto, A.; Wolstenholme, J.; Hoeksema, B.W.; et al. Relating variation in species composition to environmental variables: A multi-taxon study in an Indonesian coral reef complex. Aquat. Sci. 2008, 70, 419–431. [Google Scholar] [CrossRef]
- Huang, D.; Tun, K.P.P.; Chou, L.M.; Todd, P.A. An inventory of zooxanthellate scleractinian corals in Singapore, including 33 new records. Raffles Bull. Zool. 2009, 22, 69–80. [Google Scholar]
- Sawall, Y.; Teichberg, M.C.; Seemann, J.; Litaay, M.; Richter, C. Nutritional status and metabolism of the coral Stylophora subseriata along a eutrophication gradient in Spermonde Archipelago (Indonesia). Coral Reefs 2011, 30, 841–853. [Google Scholar] [CrossRef]
- Polónia, A.R.; Cleary, D.F.R.; de Voogd, N.J.; Renema, W.; Hoeksema, B.W.; Martins, A.; Gomes, N.C.M. Habitat and water quality variables as predictors of community composition in an Indonesian coral reef: A multi-taxon study in the Spermonde Archipelago. Sci. Total Environ. 2015, 537, 139–151. [Google Scholar] [CrossRef]
- Plass-Johnson, J.G.; Heiden, J.P.; Abu, N.; Lukman, M.; Teichberg, M. Experimental analysis of the effects of consumer exclusion on recruitment and succession of a coral reef system along a water quality gradient in the Spermonde Archipelago, Indonesia. Coral Reefs 2016, 35, 229–243. [Google Scholar] [CrossRef]
- Hopley, D. Continental shelf reef systems. In Coastal evolution, Late Quaternary Shoreline Morphodynamics; Carter, R.W.G., Woodroffe, C.D., Eds.; Cambridge University Press: Cambridge, UK, 1995; pp. 303–340. [Google Scholar]
- Janßen, A.; Wizemann, A.; Klicpera, A.; Satari, D.Y.; Westphal, H.; Mann, T. Sediment composition and facies of coral reef islands in the Spermonde Archipelago, Indonesia. Front. Mar. Sci. 2017, 4, 144. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Done, T.J. Patterns in the distribution of coral communities across the Central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Williams, D.M.B. Patterns in the distribution of fish communities across the Great Barrier Reef. Coral Reefs 1982, 1, 35–43. [Google Scholar] [CrossRef]
- Dinesen, Z.D. Patterns in the distribution of soft corals across the Central Great Barrier Reef. Coral Reefs 1983, 1, 229–236. [Google Scholar] [CrossRef]
- Preston, N.P.; Doherty, P.J. Cross-shelf patterns in the community structure of coral-dwelling Crustacea in the central region of the Great Barrier Reef. II. Cryptofauna. Mar. Ecol. Prog. Ser. 1994, 104, 27–38. [Google Scholar] [CrossRef]
- Page, C.; Willis, B. Distribution, host range and large-scale spatial variability in black band disease prevalence on the Great Barrier Reef, Australia. Dis. Aquat. Organ. 2006, 69, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becking, L.E.; Cleary, D.F.R.; de Voogd, N.J.; Renema, W.; de Beer, M.; van Soest, R.W.M.; Hoeksema, B.W. Beta-diversity of tropical marine assemblages in the Spermonde Archipelago, Indonesia. Mar. Ecol. 2006, 27, 76–88. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Distribution patterns of mushroom corals (Scleractinia:Fungiidae) across the Spermonde Shelf, Indonesia. Raffles Bull. Zool. 2012, 60, 183–212. [Google Scholar]
- Haya, L.O.M.; Fujii, M. Mapping the change of coral reefs using remote sensing and in situ measurements: A case study in Pangkajene and Kepulauan Regency, Spermonde Archipelago, Indonesia. J. Oceanogr. 2017, 73, 623–645. [Google Scholar] [CrossRef]
- Teichberg, M.; Wild, C.; Bednarz, V.N.; Kegler, H.F.; Lukman, M.; Gärdes, A.A.; Heiden, J.P.; Weiand, L.; Abu, N.; Nasir, A.; et al. Spatio-temporal patterns in coral reef communities of the Spermonde Archipelago, 2012–2014, I: Comprehensive reef monitoring of water and benthic indicators reflect changes in reef health. Front. Mar. Sci. 2018, 5, 33. [Google Scholar] [CrossRef]
- Kunzmann, A. The coral reefs of West Sumatra. In The Ecology of the Indonesian Seas; Tomasick, T., Mah, A.J., Nontji, A., Moosa, M.K., Eds.; Periplus: Singapore, 1997; Part II; pp. 1249–1262. [Google Scholar]
- Johan, O.; Prasetio, A.B.; Ardi, I.; Syam, A.R.; Quinn, N.J. The abundance of ornamental corals after mass die off in 1997 on the Padang Shelf Reef System, West Sumatera, Indonesia. J. Trop. Mar. Sci. Technol. 2016, 8, 29–38. [Google Scholar] [CrossRef]
- Renema, W. Habitat variables determining the occurrence of large benthic foraminifera in the Berau area (East Kalimantan, Indonesia). Coral Reefs 2006, 25, 351–359. [Google Scholar] [CrossRef]
- Tarya, A.; Hoitink, A.J.F.; van der Vegt, M. Tidal and subtidal flow patterns on a tropical continental shelf semi-insulated by coral reefs. J. Geophys. Res. 2010, 115, C09029. [Google Scholar] [CrossRef]
- Tarya, A.; Hoitink, A.J.F.; van der Vegt, M.; van Katwijk, M.M.; Hoeksema, B.W.; Bouma, T.J.; Lamers, L.P.M.; Christianen, M.J.A. Exposure of coastal ecosystems to river plume spreading across a near-equatorial continental shelf. Cont. Shelf Res. 2018, 153, 1–15. [Google Scholar] [CrossRef]
- Waheed, Z.; Hoeksema, B.W. A tale of two winds: Species richness patterns of reef corals around the Semporna peninsula, Malaysia. Mar. Biodivers. 2013, 43, 37–51. [Google Scholar] [CrossRef]
- DeVantier, L.; Turak, E. Coral Reefs of Brunei Darussalam; Fisheries Department, Ministry of Industry and Primary Resources, Government of Brunei Darussalam: Bandar Seri Begawan, Brunei Darussalam, 2009; pp. 1–99.
- Hoeksema, B.W.; Lane, D.J.W. The mushroom coral fauna (Scleractinia Fungiidae) of Brunei Darussalam (South China Sea) and its relation to the Coral Triangle. Raffles Bull. Zool. 2014, 62, 566–580. [Google Scholar]
- Brown, C.; Browne, N.; McIlwain, J.L.; Zinke, Z. Inshore, turbid coral reefs from northwest Borneo exhibiting low diversity, but high cover show evidence of resilience to various environmental stressors. PeerJ Preprints 2018. [Google Scholar] [CrossRef]
- Wild, C.; Jantzen, C.; Kremb, S.G. Turf algae-mediated coral damage in coastal reefs of Belize, Central America. PeerJ 2014, 2, e571. [Google Scholar] [CrossRef] [PubMed]
- Baumann, J.H.; Townsend, J.E.; Courtney, T.A.; Aichelman, H.E.; Davies, S.W.; Lima, F.P.; Castillo, K.D. Temperature regimes impact coral assemblages along environmental gradients on lagoonal reefs in Belize. PLoS ONE 2016, 11, e0162098. [Google Scholar] [CrossRef] [PubMed]
- Hoeksema, B.W. Mushroom corals (Scleractinia: Fungiidae) of Madang Lagoon, northern Papua New Guinea: An annotated checklist with the description of Cantharellus jebbi spec. nov. Zool. Meded. 1993, 67, 1–19. [Google Scholar]
- Langer, M.R.; Lipps, J.H. Foraminiferal distribution and diversity, Madang Reef and Lagoon, Papua New Guinea. Coral Reefs 2003, 22, 143–154. [Google Scholar] [CrossRef]
- Adjeroud, M.; Fernandez, J.M.; Carroll, A.G.; Harrison, P.L.; Penin, L. Spatial patterns and recruitment processes of coral assemblages among contrasting environmental conditions in the southwestern lagoon of New Caledonia. Mar. Pollut. Bull. 2010, 61, 375–386. [Google Scholar] [CrossRef]
- Gilbert, A.; Heintz, T.; Hoeksema, B.W.; Benzoni, F.; Fernandez, J.M.; Fauvelot, C.; Andréfouët, S. Endangered New Caledonian endemic mushroom coral Cantharellus noumeae in turbid, metal-rich, natural and artificial environments. Mar. Pollut. Bull. 2015, 10, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Furby, K.A.; Bouwmeester, J.; Berumen, M.L. Susceptibility of central Red Sea corals during a major bleaching event. Coral Reefs 2013, 32, 505–513. [Google Scholar] [CrossRef]
- Ellis, J.; Anlauf, H.; Kürten, S.; Lozano-Cortés, D.; Alsaffar, Z.; Cúrdia, J.; Jones, B.; Carvalho, S. Cross shelf benthic biodiversity patterns in the southern Red Sea. Sci. Rep. 2017, 7, 437. [Google Scholar] [CrossRef] [PubMed]
- Monroe, A.A.; Ziegler, M.; Roik, A.; Röthig, T.; Hardenstine, R.S.; Emms, M.A.; Jensen, T.; Voolstra, C.R.; Berumen, M.L. In situ observations of coral bleaching in the central Saudi Arabian Red Sea during the 2015/2016 global coral bleaching event. PLoS ONE 2018, 13, e0195814. [Google Scholar] [CrossRef] [PubMed]
- Umbgrove, J.H.F. De koraalriffen der Duizend Eilanden (Java-Zee). Wetensch. Meded. Dienst Mijnb. Nederl. Ind. 1929, 12, 1–47. [Google Scholar]
- Umbgrove, J.H.F. Coral reefs of the East Indies. Bull. Geol. Soc. Am. 1947, 58, 729–778. [Google Scholar] [CrossRef]
- Suyarso. Lingkungan fisik pantai dan dasar perairan Teluk Jakarta. In Atlas Oseanologi Teluk Jakarta; Suyarso, Ed.; Pusat Penelitian dan Pengembangan Oseanalogi, Lembaga Ilmu Pengetahuan Indonesia: Jakarta, Indonesia, 1995; pp. 21–27. [Google Scholar]
- Jordan, C.F. The Sedimentology of Kepulauan Seribu: A Modern Patch Reef Complex in the Java Sea, Indonesia; Indonesian Petroleum Association: Jakarta, Indonesia, 1998; pp. 1–81. [Google Scholar]
- Hoeksema, B.W. Delineation of the Indo-Malayan centre of maximum marine biodiversity: The Coral Triangle. In Biogeography, Time, and Place: Distributions, Barriers, and Islands; Renema, W., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 117–178. [Google Scholar]
- Umbgrove, J.H.F. De koraalriffen in de Baai van Batavia. Wetensch. Meded. Dienst Mijnb. Nederl. Ind. 1928, 7, 1–68. [Google Scholar]
- Umbgrove, J.H.F.; Verwey, J. The coral reefs in the Bay of Batavia. I. Geology. In Proceedings of the 4th Pacific Science Congress, Bandung, Indonesia, 16–25 May 1929; Excursion A2. pp. 1–30. [Google Scholar]
- Yates, B.F. Implementing coastal zone management policy: Kepulauan Seribu Marine National Park, Indonesia. Coast. Manag. 1994, 22, 235–249. [Google Scholar] [CrossRef]
- UNESCO. Reducing Megacity Impacts on the Coastal Environment—Alternative Livelihoods and Waste Management in Jakarta and the Seribu Islands; Coastal Region and Small Islands Paper 6; UNESCO: Paris, France, 2000; pp. 1–59. [Google Scholar]
- Fauzi, A.; Buchary, E.A. A Socioeconomic perspective of environmental degradation at Kepulauan Seribu Marine National Park, Indonesia. Coast. Manag. 2002, 30, 167–181. [Google Scholar] [CrossRef]
- Farhan, A.R.; Lim, S. Integrated vulnerability assessment on small island regions towards integrated coastal zone management (ICZM): A case study of Thousand Islands, Indonesia. Int. J. Geoinform. 2014, 10, 1–16. [Google Scholar]
- Baum, G.; Kusumanti, I.; Breckwoldt, A.; Ferse, S.C.A.; Glaser, M.; Dwiyitno; Adrianto, L.; van der Wulp, S.; Kunzmann, A. Under pressure: Investigating marine resource-based livelihoods in Jakarta Bay and the Thousand Islands. Mar. Pollut. Bull. 2016, 110, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Zaneveld, J.S.; Verstappen, H.T. A recent investigation about the geomorphology and the flora of some coral islands in the Bay of Djakarta. J. Sci. Res. 1952, 2–3, 58–68. [Google Scholar]
- Verstappen, H.T. Djakarta Bay. A Geomorphological Study on Shoreline Development. Ph.D. Thesis, Utrecht University, Utrecht, The Netherlands, 1953; pp. 1–101. [Google Scholar]
- Verstappen, H.T. Old and new observations on coastal changes of Jakarta Bay: An example of trends in urban stress on coastal environments. J. Coast. Res. 1988, 4, 573–587. [Google Scholar]
- Stoddart, D.R. Umbgrove’s islands revisited. UNESCO Rep. Mar. Sci. 1986, 40, 80–98. [Google Scholar]
- Han, Q.; Schaefer, W.; Barry, N. Land reclamation using waste as fill material: A case study in Jakarta. Int. J. Environ. Ecol. Eng. 2013, 7, 318–327. [Google Scholar]
- Van der Wulp, S.A.; Dsikowitzky, L.; Hesse, K.J.; Schwarzbauer, J. Master Plan Jakarta, Indonesia: The Giant Seawall and the need for structural treatment of municipal waste water. Mar. Pollut. Bull. 2016, 110, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Erftemeijer, P.L.A.; Riegl, B.; Hoeksema, B.W.; Todd, P.A. Environmental impacts of dredging and other sediment disturbances on corals: A review. Mar. Pollut. Bull. 2012, 64, 1737–1765. [Google Scholar] [CrossRef] [PubMed]
- Cleary, D.F.R.; Polónia, A.R.M.; Renema, W.; Hoeksema, B.W.; Wolstenholme, J.; Tuti, Y.; de Voogd, N.J. Coral reefs next to a major conurbation: A study of temporal change (1985–2011) in coral cover and composition in the reefs of Jakarta, Indonesia. Mar. Ecol. Prog. Ser. 2014, 501, 89–98. [Google Scholar] [CrossRef]
- Baum, G.; Januar, H.I.; Ferse, S.C.A.; Kunzmann, A. Local and regional impacts of pollution on coral reefs along the thousand islands north of the megacity Jakarta, Indonesia. PLoS ONE 2015, 10, e0138271. [Google Scholar] [CrossRef] [PubMed]
- Breckwoldt, A.; Dsikowitzky, L.; Baum, G.; Ferse, S.C.A.; van der Wulp, S.; Kusumanti, I.; Ramadhan, A.; Adrianto, L. A review of stressors, uses and management perspectives for the larger Jakarta Bay Area, Indonesia. Mar. Pollut. Bull. 2016, 110, 790–794. [Google Scholar] [CrossRef]
- Dsikowitzky, L.; Ferse, S.; Schwarzbauer, J.; Vogt, T.S.; Irianto, H.E. Impacts of megacities on tropical coastal ecosystems—The case of Jakarta, Indonesia. Mar. Pollut. Bull. 2016, 110, 621–623. [Google Scholar] [CrossRef] [PubMed]
- Van der Wulp, S.A.; Hesse, K.J.; Ladwig, N.; Damar, A. Numerical simulations of river discharges, nutrient flux and nutrient dispersal in Jakarta Bay, Indonesia. Mar. Pollut. Bull. 2016, 110, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Dsikowitzky, L.; Schäfer, L.; Ariyani, F.; Irianto, H.E.; Schwarzbauer, J. Evidence of massive river pollution in the tropical megacity Jakarta as indicated by faecal steroid occurrence and the seasonal flushing out into the coastal ecosystem. Environ. Chem. Lett. 2017, 15, 703–708. [Google Scholar] [CrossRef]
- Van der Meij, S.E.T.; Moolenbeek, R.G.; Hoeksema, B.W. Decline of the Jakarta Bay molluscan fauna linked to human impact. Mar. Pollut. Bull. 2009, 59, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Van der Meij, S.E.T.; Suharsono; Hoeksema, B.W. Long-term changes in coral assemblages under natural and anthropogenic stress in Jakarta Bay (1920–2005). Mar. Pollut. Bull. 2010, 60, 1442–1454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, B.E. Damage and recovery of coral reefs affected by El Niño related seawater warming in the Thousand Islands, Indonesia. Coral Reefs 1990, 8, 163–170. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Control of bleaching in mushroom coral populations (Scleractinia, Fungiidae) in the Java Sea: Stress tolerance and interference by life history strategy. Mar. Ecol. Prog. Ser. 1991, 74, 225–237. [Google Scholar] [CrossRef]
- Aziz, A.; Sukarno. Preliminary observation on living habits of Acanthaster planci (Linnaeus) at Palau Tikus, Seribu Islands. Mar. Res. Indon. 1977, 17, 121–132. [Google Scholar]
- Aziz, A. Beberapa catatan tentang kehadiran bintang laut jenis Acanthaster planci di perairan Indonesia. Oseana 1995, 20, 23–31. [Google Scholar]
- Baird, A.H.; Pratchett, M.S.; Hoey, A.S.; Herdiana, Y.; Campbell, S.J. Acanthaster planci is a major cause of coral mortality in Indonesia. Coral Reefs 2013, 32, 803–812. [Google Scholar] [CrossRef]
- Sheppard, C.R.C. Coral cover, zonation and diversity on reef slopes of Chagos Atolls, and population structures of the major species. Mar. Ecol. Prog. Ser. 1980, 2, 193–205. [Google Scholar] [CrossRef]
- Sheppard, C.R.C. The reef and soft-substrate coral fauna of Chagos, Indian Ocean. J. Nat. Hist. 1981, 15, 607–621. [Google Scholar] [CrossRef]
- Graus, R.R.; Macintyre, I.G. The zonation patterns of Caribbean coral reefs as controlled by wave and light energy input, bathymetric setting and reef morphology: Computer simulation experiments. Coral Reefs 1989, 8, 9–18. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Moka, W. Species assemblages and phenotypes of mushroom corals (Scleractinia: Fungiidae) related to coral reef habitats in the Flores Sea. Neth. J. Sea Res. 1989, 23, 149–160. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Evolutionary trends in onshore-offshore distribution patterns of mushroom coral species (Scleractinia: Fungiidae). Contrib. Zool. 2012, 81, 199–221. [Google Scholar]
- Lane, D.J.W.; Hoeksema, B.W. Mesophotic mushroom coral records at Brunei Darussalam support westward extension of the Coral Triangle to the South China Sea waters of Northwest Borneo. Raffles Bull. Zool. 2016, 64, 566–580. [Google Scholar]
- Hoeksema, B.W. Latitudinal species diversity gradient of mushroom corals off eastern Australia: A baseline from the 1970s. Estuar. Coast. Shelf Sci. 2015, 165, 190–198. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Sellanes, J.; Easton, E.E. A high-latitude, mesophotic Cycloseris field at 85 m depth off Rapa Nui (Easter Island). Bull. Mar. Sci. 2019, 95, 101–102. [Google Scholar] [CrossRef]
- Goreau, T.F.; Yonge, C.M. Coral community on muddy sand. Nature 1968, 217, 421–423. [Google Scholar] [CrossRef]
- Pichon, M. Free living scleractinian coral communities in the coral reefs of Madagascar. In Proceedings of the 2nd International Coral Reef Symposium, Brisbane, Australian, 2 June–22 July 1974; Volume 2, pp. 173–181. [Google Scholar]
- Fisk, D.A. Free-living corals: Distributions according to plant cover, sediments, hydrodynamics, depth and biological factors. Mar. Biol. 1983, 74, 287–294. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Cairns, S. World List of Scleractinia. Accessed through: World Register of Marine Species. Available online: http://www.marinespecies.org/scleractinia/ (accessed on 31 January 2019).
- Hoeksema, B.W. Attached mushroom corals (Scleractinia: Fungiidae) in sediment-stressed reef conditions at Singapore, including a new species and a new record. Raffles Bull. Zool. 2009, Suppl. 22, 81–90. [Google Scholar]
- Benzoni, F.; Arrigoni, R.; Stefani, F.; Reijnen, B.T.; Montano, S.; Hoeksema, B.W. Phylogenetic position and taxonomy of Cycloseris explanulata and C. wellsi (Scleractinia: Fungiidae): Lost mushroom corals find their way home. Contrib. Zool. 2012, 81, 125–146. [Google Scholar]
- Hoeksema, B.W. Evolution of body size in mushroom corals (Scleractinia: Fungiidae) and its ecomorphological consequences. Neth. J. Zool. 1991, 41, 122–139. [Google Scholar] [CrossRef]
- Gittenberger, A.; Reijnen, B.T.; Hoeksema, B.W. A molecularly based phylogeny reconstruction of mushroom corals (Scleractinia: Fungiidae) with taxonomic consequences and evolutionary implications for life history traits. Contrib. Zool. 2011, 80, 107–132. [Google Scholar]
- Hoeksema, B.W. The “Fungia patella group” (Scleractinia, Fungiidae) revisited with a description of the mini mushroom coral Cycloseris boschmai sp. n. ZooKeys 2014, 371, 57–84. [Google Scholar] [CrossRef]
- Umbgrove, J.H.F. Madreporaria from the Bay of Batavia. Zool. Meded. 1939, 22, 1–64. [Google Scholar]
- Moll, H.; Suharsono. Distribution, diversity and abundance of reef corals in Jakarta Bay and Kepulauan Seribu. UNESCO Rep. Mar. Sci. 1986, 40, 112–125. [Google Scholar]
- Soemodihardjo, S. (Ed.) Proceedings: Coral Reef Evaluation Workshop Pulau Seribu, Jakarta, Indonesia, 11–20 September 1995; Contending with Global Change 10; UNESCO: Jakarta, Indonesia, 1998; pp. 1–129. [Google Scholar]
- Hoeksema, B.W. Taxonomy, phylogeny and biogeography of mushroom corals (Scleractinia: Fungiidae). Zool. Verh. 1989, 254, 1–295. [Google Scholar]
- Moll, H. Zonation and Diversity of Scleractinia on Reefs off S.W. Sulawesi, Indonesia. Ph.D. Thesis, Leiden University, Leiden, The Netherlands, 1983. [Google Scholar]
- Schuhmacher, H.; Mergner, H. Quantitative Analyse von Korallengemeinschaften des Sanganeb-Atolls (mittleres Rotes Meer). II. Vergleich mit einem Riffareal bei Aqaba (nördliches Rotes Meer) am Nordrande des indopazifischen Riffgürtels. Helgol. Meeresunters. 1985, 39, 419–440. [Google Scholar] [CrossRef] [Green Version]
- Waheed, Z.; Hoeksema, B.W. Diversity patterns of scleractinian corals at Kota Kinabalu, Malaysia, in relation to exposure and depth. Raffles Bull. Zool. 2014, 62, 66–82. [Google Scholar]
- Waheed, Z.; van Mil, H.G.J.; Syed Hussein, M.A.; Jumin, R.; Golam Ahad, B.; Hoeksema, B.W. Coral reefs at the northernmost tip of Borneo: An assessment of scleractinian species richness patterns and benthic reef assemblages. PLoS ONE 2015, 10, e0146006. [Google Scholar] [CrossRef]
- Muir, P.R.; Wallace, C.C.; Bridge, T.C.L.; Bongaerts, P. Diverse staghorn coral fauna on the mesophotic reefs of North-East Australia. PLoS ONE 2015, 10, e0117933. [Google Scholar] [CrossRef] [PubMed]
- Muir, P.R.; Wallace, C.C.; Pichon, M.; Bongaerts, P. High species richness and lineage diversity of reef corals in the mesophotic zone. Proc. R. Soc. B 2018, 285, 20181987. [Google Scholar] [CrossRef]
- Muir, P.R.; Pichon, M.; Squire, L.; Wallace, C.C. Acropora tenella, a zooxanthellate coral extending to 110-m depth in the northern Coral Sea. Mar. Biodivers. 2018. [Google Scholar] [CrossRef]
- Larcombe, P.; Woolfe, K.J. Increased sediment supply to the Great Barrier Reef will not increase sediment accumulation at most coral reefs. Coral Reefs 1999, 18, 163–169. [Google Scholar] [CrossRef]
- Larcombe, P.; Costen, A.; Woolfe, K.J. The hydrodynamic and sedimentary setting of nearshore coral reefs, central Great Barrier Reef shelf, Australia: Paluma Shoals, a case study. Sedimentology 2001, 48, 811–835. [Google Scholar] [CrossRef]
- Tuti, M.I.Y.; Soemodihardjo, S. (Eds.) Coral Reef Ecosystem in the Seribu Islands; LIPI Press: Jakarta, Indonesia, 2006; pp. 1–179. [Google Scholar]
- Brown, B.E. (Ed.) Human Induced Damage to Coral Reefs. UNESCO Rep. Mar. Sci. 1986, 40, 1–180. [Google Scholar]
- Cleary, D.F.R.; Suharsono; Hoeksema, B.W. Coral diversity across a disturbance gradient in the Pulau Seribu reef complex off Jakarta, Indonesia. Biodivers. Conserv. 2006, 15, 3653–3674. [Google Scholar] [CrossRef]
- TNI-AL. Pulau-Pulau Seribu. Pulau Rambut Hingga Karang Jalan, Sekala 1:20.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1997; 410-KK. [Google Scholar]
- TNI-AL. Teluk Jakarta. Pulau Rambut Hingga Pelabuhan Tanjungpriok, Sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1996; 411-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Karang Tohorjantan Hingga Pulau Damar Besar, Sekala 1:50.000; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1985; 412-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Tidung Besar Hingga Pulau Bokor, Sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1998; 413-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Jukung Hingga Pulau Piniki, Sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1995; 414-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Dua Barat hingga Pulau Tondan Timur, sekala 1:50.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1998; 415-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Pramuka hingga Pulau Kotok Kecil, sekala 1:20.000; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1986; 416-KK. [Google Scholar]
- TNI-AL. Pulau-Pulau Seribu. Pulau Opak Kecil hingga Pulau Jukung, sekala 1:20.000, 2nd ed.; Tentara Nasional Indonesia—Ankatan Laut, Jawatan Hidro-Oseanografi: Jakarta, Indonesia, 1998; 417-KK. [Google Scholar]
- English, S.; Wilkinson, C.; Baker, V. Survey Manual for Tropical Marine Resources, 2nd ed.; Australian Institute of Marine Science: Townsville, Australia, 1997; pp. 1–390. [Google Scholar]
- Google Maps. Available online: https://www.google.com/maps/ (accessed on 12 January 2019).
- GPS Coordinates. Available online: https://gps-coordinates.org (accessed on 16 January 2019).
- Giyanto; Tuti, Y.; Budiyanto, A. Preliminary analysis on the condition of coral reefs in the Kepulaan Seribu, Jakarta 2005. In Coral Reef Ecosystem in the Seribu Islands: Workshop Monitoring and Evaluation of Three Decades; Tuti, Y., Soemodihardjo, S., Eds.; LIPI Press: Jakarta, Indonesia, 2006; pp. 9–17. [Google Scholar]
- Hoeksema, B.W.; Koh, E.G.L. Depauperation of the mushroom coral fauna (Fungiidae) of Singapore (1860s–2006) in changing reef conditions. Raffles Bull. Zool. 2009, 22, 91–101. [Google Scholar]
- Veron, J.E.N. Corals of the World; Australian Institute of Marine Science: Townsville, Australia, 2000; Volume 2, pp. 1–429. [Google Scholar]
- Benzoni, F.; Stefani, F.; Stolarski, J.; Pichon, M.; Mitta, G.; Galli, P. Debating phylogenetic relationships of the scleractinian Psammocora: Molecular and morphological evidences. Contrib. Zool. 2007, 76, 35–54. [Google Scholar]
- MINITAB. Minitab 17.1.0; Minitab Inc.: State College, PA, USA, 2013. [Google Scholar]
- Colwell, R.K. EstimateS: Statistical Estimation of Species Richness and Shared Species from Samples, Version 9.1; User’s Guide and Application. 2013. Available online: http://viceroy.eeb.uconn.edu/estimates (accessed on 20 December 2018).
- Verwey, J. Coral reef studies. II. The depth of coral reefs in relation to their oxygen consumption and the penetration of light in the water. Treubia 1931, 13, 169–198. [Google Scholar]
- Harger, J.R.E. Community displacement in stressed coral reef systems and the implication for a comprehensive management strategy for coastal and offshore productivity enrichment. Galaxea 1988, 7, 185–196. [Google Scholar]
- Tomascik, T.; Suharsono; Mah, A.J. Case histories: A historical perspective of the natural and anthropogenic impacts in the Indonesian archipelago with a focus on Kepulauan Seribu, Java Sea. In Proceedings of the Colloquium on Global Aspects of Coral Reefs, Health, Hazards and History; Ginsburg, R., Ed.; University of Miami, Rosenstiel School of Marine and Atmospheric Science: Miami, FL, USA, 1993; pp. 304–310. [Google Scholar]
- Johan, O.; Zamany, N.P.; Smith, D.; Sweet, M.J. Prevalence and incidence of black band disease of scleractinian corals in the Kepulauan Seribu Region of Indonesia. Diversity 2016, 8, 11. [Google Scholar] [CrossRef]
- Baum, G.; Januar, H.I.; Ferse, S.C.A.; Wild, C.; Kunzmann, A. Abundance and physiology of dominant soft corals linked to water quality in Jakarta Bay, Indonesia. PeerJ 2016, 4, e2625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleary, D.F.R.; Polónia, A.R.M.; Renema, W.; Hoeksema, B.W.; Rachello-Dolmen, P.G.; Moolenbeek, R.G.; Budiyanto, A.; Yahmantoro; Tuti, Y.; Draisma, S.G.A.; et al. Variation in the composition of corals, fishes, sponges, echinoderms, ascidians, molluscs, foraminifera and macroalgae across a pronounced in-to-offshore environmental gradient in the Jakarta Bay–Thousand Islands coral reef complex. Mar. Pollut. Bull. 2016, 110, 701–717. [Google Scholar] [CrossRef]
- Draisma, S.G.A.; Prud'homme van Reine, W.F.; Herandarudewi, S.M.C.; Hoeksema, B.W. Macroalgal diversity along an inshore-offshore environmental gradient in the Jakarta Bay–Thousand Islands reef complex, Indonesia. Estuar. Coast. Shelf Sci. 2018, 200, 258–269. [Google Scholar] [CrossRef]
- Madduppa, H.H.; Subhan, B.; Suparyani, E.; Siregar, A.M.; Arafat, D.; Tarigan, S.A.; Alimuddin; Khairudi, D.; Rahmawati, F.; Bramandito, A. Dynamics of fish diversity across an environmental gradient in the Seribu Islands reefs off Jakarta. Biodiversitas 2013, 14, 17–24. [Google Scholar] [CrossRef]
- De Voogd, N.J.; Cleary, D.F.R. An analysis of sponge diversity and distribution at three taxonomic levels in the Thousand Islands/Jakarta bay reef complex, West-Java, Indonesia. Mar. Ecol. 2008, 29, 205–215. [Google Scholar] [CrossRef]
- Siregar, T.H.; Priyanto, N.; Putri, A.K.; Rachmawati, N.; Triwibowo, R.; Dsikowitzky, L.; Schwarzbauer, J. Spatial distribution and seasonal variation of the trace hazardous element contamination in Jakarta Bay, Indonesia. Mar. Pollut. Bull. 2016, 110, 634–646. [Google Scholar] [CrossRef]
- Alder, J.; Sloan, N.A.; Uktolseya, H.A. comparison of management planning and implementation in three Indonesian marine protected areas. Ocean Coast. Manag. 1994, 24, 179–198. [Google Scholar] [CrossRef]
- Farhan, A.R.; Lim, S. Vulnerability assessment of ecological conditions in Seribu Islands, Indonesia. Ocean Coast. Manag. 2012, 65, 1–14. [Google Scholar] [CrossRef]
- Erdmann, M.V. Destructive fishing practices in the Pulau Seribu Archipelego. In Proceedings of the Coral Reef evaluation workshop Pulau Seribu, Jakarta, Indonesia, 11–20 September 1995; Soemodihardjo, S., Ed.; Contending with Global Change 10. UNESCO: Jakarta, Indonesia, 1998; pp. 84–89. [Google Scholar]
- Fox, H.E.; Caldwell, R.L. Recovery from blast fishing on coral reefs: A tale of two scales. Ecol. Appl. 2006, 16, 1631–1635. [Google Scholar] [CrossRef]
- Raymundo, L.J.; Maypa, A.P.; Gomez, E.D.; Cadiz, P. Can dynamite-blasted reefs recover? A novel, low-tech approach to stimulating natural recovery in fish and coral populations. Mar. Pollut. Bull. 2007, 54, 1009–1019. [Google Scholar] [CrossRef]
- Williams, S.L.; Sur, C.; Janetski, N.; Hollarsmith, J.A.; Rapi, S.; Barron, L.; Heatwole, S.J.; Yusuf, A.M.; Yusuf, S.; Jompa, J.; et al. Large-scale coral reef rehabilitation after blast fishing in Indonesia. Restor. Ecol. 2018. [Google Scholar] [CrossRef]
- Fox, H.E.; Harris, J.L.; Darling, E.S.; Ahmadia, G.N.; Estradivari; Razak, T.B. Rebuilding coral reefs: Success (and failure) 16 years after low-cost, low-tech restoration. Restor. Ecol. 2019. [Google Scholar] [CrossRef]
- Hoeksema, B.W. Mobility of free-living fungiid corals (Scleractinia), a dispersion mechanism and survival strategy in dynamic reef habitats. In Proceedings of the 6th International Coral Reef Symposium, Townsville, Australia, 8–12 August 1988; Volume 2, pp. 715–720. [Google Scholar]
- Best, M.B.; Hoeksema, B.W.; Moka, W.; Moll, H.; Suharsono; Sutarna, I.N. Recent scleractinian coral species collected during the Snellius-II Expedition in eastern Indonesia. Neth. J. Sea Res. 1989, 23, 107–115. [Google Scholar] [CrossRef]
- Huang, D.; Licuanan, W.Y.; Hoeksema, B.W.; Chen, C.A.; Ang, P.O.; Huang, H.; Lane, D.J.W.; Vo, S.T.; Waheed, Z.; Amri, A.Y.; et al. Extraordinary diversity of reef corals in the South China Sea. Mar. Biodivers. 2015, 45, 157–168. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Waheed, Z. Initial phase of autotomy in fragmenting Cycloseris corals at Semporna, eastern Sabah, Malaysia. Coral Reefs 2011, 30, 1087. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Bouwmeester, J.; Range, P.; Ben-Hamadou, R.A. large aggregation of self-fragmenting mushroom corals in the Arabian/Persian Gulf. Ecology 2018, 99, 1236–1238. [Google Scholar] [CrossRef] [PubMed]
- Schuhmacher, H. Ability in fungiid corals to overcome sedimentation. In Proceedings of the 3rd International Coral Reef Symposium, Miami, 1977; pp. 503–509. [Google Scholar]
- Hoeksema, B.W.; Yeemin, T. Late detachment conceals serial budding by the free-living coral Fungia fungites in the Inner Gulf of Thailand. Coral Reefs 2011, 30, 975. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Waheed, Z. Onset of autotomy in an attached Cycloseris coral. Galaxea J. Coral Reef Stud. 2012, 14, 1–2. [Google Scholar] [CrossRef]
- Bongaerts, P.; Hoeksema, B.W.; Hay, K.B.; Hoegh-Guldberg, O. Mushroom corals overcome live burial through pulsed inflation. Coral Reefs 2012, 31, 399. [Google Scholar] [CrossRef]
- Chadwick-Furman, N.E.; Loya, Y. Migration, habitat use, and competition among mobile corals (Scleractinia: Fungiidae) in the Gulf of Eilat, Red Sea. Mar. Biol. 1992, 114, 617–623. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; de Voogd, N.J. On the run: Free-living mushroom corals avoiding interaction with sponges. Coral Reefs 2012, 31, 455–459. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Bongaerts, P. Mobility and self-righting by a freeliving mushroom coral through pulsed inflation. Mar. Biodivers. 2016, 46, 521–524. [Google Scholar] [CrossRef]
- Plusquellec, Y.; Webb, G.; Hoeksema, B.W. Automobility in Tabulata, Rugosa, and extant scleractinian analogues: Stratigraphic and paleogeographic distribution of Paleozoic mobile corals. J. Paleontol. 1999, 73, 985–1001. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Hassell, D.; Meesters, E.H.W.G.; van Duyl, F.C. Wave-swept coralliths of Saba Bank, Dutch Caribbean. Mar. Biodivers. 2018, 48, 2003–2016. [Google Scholar] [CrossRef]
- Denis, V.; De Palmas, S.; Benzoni, F.; Chen, C.C. Extension of the known distribution and depth range of the scleractinian coral Psammocora stellata: First record from a Taiwanese mesophotic reef. Mar. Biodivers. 2015, 45, 619–620. [Google Scholar] [CrossRef]
Figure 1.
Map of Jakarta Bay and the Thousand Islands archipelago (Kepulauan Seribu) off Jakarta. Reefs surveyed in 2005 (Table 1) are indicated by their name in different colors to distinguish three shelf zones that vary in distance offshore and depth range: Red = Zone 1 (0–30 m), yellow = Zone 2 (30–60 m), blue = Zone 3 (20–40 m). The reference point for Tanjung Priok harbor is marked.
Figure 1.
Map of Jakarta Bay and the Thousand Islands archipelago (Kepulauan Seribu) off Jakarta. Reefs surveyed in 2005 (Table 1) are indicated by their name in different colors to distinguish three shelf zones that vary in distance offshore and depth range: Red = Zone 1 (0–30 m), yellow = Zone 2 (30–60 m), blue = Zone 3 (20–40 m). The reference point for Tanjung Priok harbor is marked.

Figure 2.
Significant relations between abiotic parameters off Jakarta: (a) Shelf depth in relation to latitude following quadratic model; (b) water transparency (secchi depth) in relation to distance from mainland, quadratic model; (c) secchi depth in relation to shelf depth, linear model. Color codes correspond with those of zones in Figure 1: Zone 1 (red), Zone 2 (yellow), Zone 3 (blue).
Figure 2.
Significant relations between abiotic parameters off Jakarta: (a) Shelf depth in relation to latitude following quadratic model; (b) water transparency (secchi depth) in relation to distance from mainland, quadratic model; (c) secchi depth in relation to shelf depth, linear model. Color codes correspond with those of zones in Figure 1: Zone 1 (red), Zone 2 (yellow), Zone 3 (blue).

Figure 3.
Significant relations (quadratic model) between mushroom coral diversity and abiotic parameters off Jakarta (2–30 m depth range): (a) shelf depth; (b) secchi depth; (c) latitude; (d) distance from mainland; (e) distance from harbor; (f) distance to nearest river outlet. Color codes correspond with those of zones in Figure 1: Zone 1 (red), Zone 2 (yellow), Zone 3 (blue).
Figure 3.
Significant relations (quadratic model) between mushroom coral diversity and abiotic parameters off Jakarta (2–30 m depth range): (a) shelf depth; (b) secchi depth; (c) latitude; (d) distance from mainland; (e) distance from harbor; (f) distance to nearest river outlet. Color codes correspond with those of zones in Figure 1: Zone 1 (red), Zone 2 (yellow), Zone 3 (blue).

Figure 4.
Shallow-water mushroom corals in zones 2–3: (a) Halomitra pileus at NW Pulau Peniki; (b) Lobactis scutaria at W Pulau Tikus; (c) Pleuractis granulosa at NW Pulau Kotok Kecil; (d) attached coral of Podabacia kunzmanni from N Pulau Panjang, 4 m depth (RMNH Coel. 24176, 18.ix.1995, coll. B.W.H.); (e) oral side of Lithophyllon scabra corallum from Pulau Tidung Besar (RMNH Coel. 16101, 12.v.1983, coll. B.W.H.); (f) aboral side of the same specimen.
Figure 4.
Shallow-water mushroom corals in zones 2–3: (a) Halomitra pileus at NW Pulau Peniki; (b) Lobactis scutaria at W Pulau Tikus; (c) Pleuractis granulosa at NW Pulau Kotok Kecil; (d) attached coral of Podabacia kunzmanni from N Pulau Panjang, 4 m depth (RMNH Coel. 24176, 18.ix.1995, coll. B.W.H.); (e) oral side of Lithophyllon scabra corallum from Pulau Tidung Besar (RMNH Coel. 16101, 12.v.1983, coll. B.W.H.); (f) aboral side of the same specimen.

Figure 5.
Free-living mid-slope mushroom corals at zones 2–3: (a) Ctenactis albitentaculata at NW Pulau Belanda; (b) Ctenactis crassa at S Pulau Tikus; (c) Cycloseris boschmai at E Karang Jong; (d) C. cyclolites at S Pulau Pari; (e) C. tenuis at E Pulau Peniki; (f) Sandalolitha dentata at S Pulau Kotok Kecil.
Figure 5.
Free-living mid-slope mushroom corals at zones 2–3: (a) Ctenactis albitentaculata at NW Pulau Belanda; (b) Ctenactis crassa at S Pulau Tikus; (c) Cycloseris boschmai at E Karang Jong; (d) C. cyclolites at S Pulau Pari; (e) C. tenuis at E Pulau Peniki; (f) Sandalolitha dentata at S Pulau Kotok Kecil.

Figure 6.
Cycloseris corals, encrusting on lower-slopes (a,b) and free-living on reef-bases (c–f) in zones 2–3: (a) C. explanulata at E Pulau Pari; (b) C. mokai at NW Pulau Hantu Kecil; (c) C. fragilis (self-fragmenting) at NW Pulau Peniki; (d) C. fragilis (complete) at NW Pulau Tidung Besar; (e) C. sinensis (self-fragmenting) at NW Pulau Peniki; (f) C. vaughani at E Karang Jong.
Figure 6.
Cycloseris corals, encrusting on lower-slopes (a,b) and free-living on reef-bases (c–f) in zones 2–3: (a) C. explanulata at E Pulau Pari; (b) C. mokai at NW Pulau Hantu Kecil; (c) C. fragilis (self-fragmenting) at NW Pulau Peniki; (d) C. fragilis (complete) at NW Pulau Tidung Besar; (e) C. sinensis (self-fragmenting) at NW Pulau Peniki; (f) C. vaughani at E Karang Jong.

Figure 7.
Significant relations between mushroom coral diversity and abiotic parameters off Jakarta for shallow reef depths (2–6 m): (a) shelf depth; (b) secchi depth; (c) latitude.
Figure 7.
Significant relations between mushroom coral diversity and abiotic parameters off Jakarta for shallow reef depths (2–6 m): (a) shelf depth; (b) secchi depth; (c) latitude.

Figure 8.
Reef flat of Pulau Putri at ca. 5 m depth (Zone 3), damaged by blast fishing. Free-living mushroom corals (arrows) are scattered among dead and live fragments of other corals: (a,b) Fungia fungites; (c,d) Lithophyllon repanda. Date: 14.ix.2005.
Figure 8.
Reef flat of Pulau Putri at ca. 5 m depth (Zone 3), damaged by blast fishing. Free-living mushroom corals (arrows) are scattered among dead and live fragments of other corals: (a,b) Fungia fungites; (c,d) Lithophyllon repanda. Date: 14.ix.2005.

Figure 9.
Species richness estimators covering 29 mushroom coral species observed in 38 reef sites based on incidence data.
Figure 9.
Species richness estimators covering 29 mushroom coral species observed in 38 reef sites based on incidence data.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Reef sites off Jakarta surveyed for Fungiidae with coordinates; shelf zone (Figure 1); shelf depth; secchi depth; and shortest distance to mainland, river outlet, and Tanjung Priok harbor (coordinates −6.0932, 106.8770).
Table 1.
Reef sites off Jakarta surveyed for Fungiidae with coordinates; shelf zone (Figure 1); shelf depth; secchi depth; and shortest distance to mainland, river outlet, and Tanjung Priok harbor (coordinates −6.0932, 106.8770).
| Site | Coordinates, DD | Shelf Zone | Number of Species | Shallow Records | Shelf Depth, m | Secchi Depth, m | Nearest Mainland, km | Nearest River Outlet, km | Distance to Harbor, km |
|---|---|---|---|---|---|---|---|---|---|
| Onrust NW | −6.0334, 106.7328 | 1 | 0 | 0 | 9 | 1.5 | 2.3 | 2.3 | 17.3 |
| Bidadari NW | −6.0321, 106.7463 | 1 | 5 | 5 | 11 | 1.5 | 3.6 | 3.7 | 16.0 |
| Nyamuk Besar NW | −6.0291, 106.8523 | 1 | 2 | 2 | 18 | 3.5 | 7.6 | 12.9 | 7.6 |
| Kelor NW | −6.0253, 106.7441 | 1 | 3 | 3 | 12 | 2.5 | 3.8 | 3.8 | 16.5 |
| Ayer Besar NW | −6.0017, 106.7801 | 1 | 1 | 1 | 18 | 3.5 | 8.6 | 8.6 | 14.8 |
| Ubi Besar NW | −5.9987, 106.7397 | 1 | 3 | 2 | 15 | 4.0 | 5.5 | 5.8 | 18.5 |
| Damar Kecil NW | −5.9834, 106.8453 | 1 | 5 | 4 | 19 | 3.5 | 12.7 | 15.9 | 12.7 |
| Untung Jawa NW | −5.9741, 106.7031 | 1 | 10 | 9 | 20 | 2.5 | 5.0 | 6.6 | 23.4 |
| Damar Besar NW | −5.8547, 106.8409 | 1 | 10 | 10 | 28 | 8.0 | 15.4 | 15.4 | 15.9 |
| Dapur NW | −5.9457, 106.7242 | 2 | 17 | 14 | 28 | 7.5 | 8.9 | 10.3 | 23.6 |
| Bokor NW | −5.9430, 106.6271 | 1 | 10 | 9 | 20 | 5.0 | 6.7 | 6.7 | 32.2 |
| Lancang NW | −5.9270, 106.5913 | 1 | 5 | 5 | 21 | 5.0 | 9.5 | 9.5 | 36.6 |
| Pari S shoal | −5.8808, 106.6352 | 2 | 15 | - | 29 | 8.0 | 12.0 | 13.6 | 35.7 |
| Tikus S | −5.8653, 106.5819 | 2 | 15 | 10 | 32 | 6.5 | 15.5 | 16.3 | 41.3 |
| Tikus W | −5.8589, 106.5700 | 2 | 14 | 9 | 37 | 8.0 | 16.8 | 17.4 | 42.8 |
| Pari E | −5.8544, 106.6389 | 2 | 17 | 8 | 37 | 12.5 | 14.8 | 16.6 | 37.4 |
| Tikus N | −5.8536, 106.5786 | 2 | 17 | 12 | 43 | 11.0 | 16.9 | 17.6 | 42.4 |
| Karang Jong E | −5.8522, 106.6486 | 2 | 20 | 11 | 34 | 11.5 | 15.0 | 16.9 | 36.8 |
| Karang Jong NW | −5.8511, 106.6464 | 2 | 19 | 12 | 48 | 11.0 | 15.1 | 17.0 | 37.1 |
| Tikus NE | −5.8501, 106.5848 | 2 | 18 | 11 | 34 | 8.5 | 16.9 | 17.8 | 42.1 |
| Payung Besar E | −5.8219, 106.5631 | 2 | 15 | 11 | 54 | 10.5 | 19.4 | 21.5 | 46.0 |
| Payung Kecil NW | −5.8134, 106.5492 | 2 | 19 | 11 | 54 | 15.5 | 20.1 | 23.0 | 47.8 |
| Tidung Kecil NW | −5.7997, 106.5178 | 2 | 21 | 13 | 45 | 17.5 | 21.7 | 25.9 | 51.4 |
| Tidung Besar NW | −5.7910, 106.4812 | 2 | 17 | 10 | 43 | 15.0 | 23.3 | 28.8 | 55.2 |
| Air NW | −5.7606, 106.7456 | 3 | 15 | 11 | 29 | 12.0 | 27.3 | 28.9 | 39.7 |
| Semak Daun NW | −5.7322, 106.5731 | 3 | 16 | 11 | 28 | 11.5 | 29.6 | 30.8 | 52.4 |
| Kotok Besar NW | −5.6988, 106.5398 | 3 | 16 | 12 | 33 | 10.5 | 32.8 | 35.3 | 57.6 |
| Peniki NW | −5.6968, 106,7155 | 2 | 23 | 12 | 43 | 9.5 | 32.3 | 34.9 | 47.6 |
| Peniki E | −5.6922, 106.7174 | 2 | 21 | 11 | 41 | 9.5 | 33.6 | 35.4 | 48.0 |
| Kotok Kecil NW | −5.6888, 106.5336 | 3 | 19 | 15 | 25 | 10.0 | 33.9 | 36.6 | 58.9 |
| Kelapa, NW | −5.6544, 106.5589 | 3 | 15 | 13 | 28 | 11.0 | 37.8 | 39.6 | 60.2 |
| Panjang, NW | −5.6423, 106.5599 | 3 | 14 | 12 | 26 | 11.5 | 39.2 | 40.9 | 61.2 |
| Belanda, NW | −5.6037, 106.6035 | 3 | 19 | 12 | 31 | 10.0 | 43.0 | 44.5 | 62.3 |
| Putri, NW | −5.5904, 106.5673 | 3 | 15 | 11 | 26 | 10.5 | 45.0 | 46.4 | 65.6 |
| Sepa, NW | −5.5755, 106.5799 | 3 | 15 | 13 | 29 | 11.0 | 46.4 | 47.9 | 66.3 |
| Jukung, NW | −5.5669, 106.5272 | 3 | 11 | 10 | 30 | 9.5 | 47.5 | 49.9 | 70.2 |
| Hantu Kecil, NW | −5.5354, 106.5319 | 3 | 19 | 15 | 31 | 10.0 | 51.0 | 53.2 | 72.3 |
| Hantu Besar, NW | −5.5296, 106.5389 | 3 | 17 | 14 | 27 | 9.5 | 51.6 | 53.6 | 73.0 |
Table 2.
Numbers of mushroom coral species (n = 31) encountered off Jakarta: 29 species recorded in 2005 and two additional ones observed in 1983 and 1995; # = new species record for the area. Shallow-water records (if any) are indicated by brackets.
Table 2.
Numbers of mushroom coral species (n = 31) encountered off Jakarta: 29 species recorded in 2005 and two additional ones observed in 1983 and 1995; # = new species record for the area. Shallow-water records (if any) are indicated by brackets.
| Species | Zone 1 11 Sites | Zone 2 15 Sites | Zone 3 12 Sites | All Three Zones 38 Sites |
|---|---|---|---|---|
| Ctenactis albitentaculata (Hoeksema, 1989) | 1 (1) | 10 (2) | 8 (5) | 19 (8) |
| Ctenactis crassa (Dana, 1846) | 8 | 9 (6) | 17 (6) | |
| Ctenactis echinata (Pallas, 1766) | 4 (4) | 14 (13) | 12 (12) | 30 (29) |
| Cycloseris boschmai (Hoeksema, 2014) # | 2 | 2 | ||
| Cycloseris costulata (Ortmann, 1889) | 3 (2) | 11 | 1 | 15 (2) |
| Cycloseris cyclolites (Lamarck, 1816) # | 3 | 3 | ||
| Cycloseris explanulata (van der Horst, 1922) # | 3 | 1 | 4 | |
| Cycloseris fragilis (Alcock, 1893) | 10 | 1 | 11 | |
| Cycloseris mokai (Hoeksema, 1989) | 7 | 4 | 11 | |
| Cycloseris sinensis (Milne Edwards and Haime, 1851) # | 1 | 2 | 3 | |
| Cycloseris tenuis (Dana, 1846) # | 1 | 1 | ||
| Cycloseris vaughani (Boschma, 1923) # | 1 | 1 | ||
| Danafungia horrida (Dana, 1846) | 2 (2) | 15 (14) | 12 (12) | 29 (28) |
| Danafungia scruposa (Klunzinger, 1879) | 2 (2) | 13 (12) | 11 (11) | 26 (25) |
| Fungia fungites (Linnaeus, 1758) | 6 (6) | 15 (14) | 12 (12) | 33 (32) |
| Halomitra pileus (Linnaeus, 1758) | 2 (2) | 2 (1) | 4 (3) | |
| Heliofungia actiniformis (Quoy and Gaimard, 1833) | 2 (2) | 10 (2) | 8 (6) | 20 (10) |
| Herpolitha limax (Esper, 1797) | 3 (1) | 15 (14) | 12 (12) | 30 (27) |
| Lithophyllon concinna (Verrill, 1864) | 2 (2) | 15 (14) | 12 (12) | 29 (28) |
| Lithophyllon repanda (Dana, 1846) | 3 (3) | 15 (14) | 12 (12) | 30 (29) |
| Lithophyllon scabra (Döderlein, 1901) | 1 * | 1 * | ||
| Lithophyllon undulatum Rehberg, 1892 | 5 (5) | 7 (1) | 5 (1) | 17 (7) |
| Lobactis scutaria (Lamarck, 1801) | 3 (1) | 3 (1) | ||
| Pleuractis granulosa (Klunzinger, 1879) | 10 (5) | 6 (5) | 16 (10) | |
| Pleuractis moluccensis (van der Horst, 1919) | 3 (2) | 15 (1) | 10 | 28 (3) |
| Pleuractis paumotensis (Stutchbury, 1833) | 5 (5) | 15 (14) | 12 (12) | 32 (31) |
| Podabacia crustacea (Pallas, 1766) | 1 (1) | 13 (8) | 12 (12) | 26 (21) |
| Podabacia kunzmanni (Hoeksema, 2009) # | (1)** | (1)** | ||
| Polyphyllia talpina (Lamarck, 1801) | 4 (4) | 11 (11) | 7 (7) | 22 (22) |
| Sandalolitha dentata Quelch, 1884 | 9 | 8 (1) | 17 (1) | |
| Sandalolitha robusta (Quelch, 1886) | 8 (8) | 14 (12) | 12 (11) | 34 (31) |
| Total number of species per zone/three zones | 16 (16) | 29 (18) + 1 * | 24 (18) + 1 ** | 29 (21) + 2 |
* Record from Pulau Payung Besar in 1983; ** record from Pulau Panjang in 1995.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Hoeksema, B.W.; Giyanto; Suharsono. The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta. Diversity 2019, 11, 46. https://doi.org/10.3390/d11030046
AMA Style
Hoeksema BW, Giyanto, Suharsono. The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta. Diversity. 2019; 11(3):46. https://doi.org/10.3390/d11030046
Chicago/Turabian StyleHoeksema, Bert W., Giyanto, and Suharsono. 2019. "The Role of Maximum Shelf Depth versus Distance from Shore in Explaining a Diversity Gradient of Mushroom Corals (Fungiidae) off Jakarta" Diversity 11, no. 3: 46. https://doi.org/10.3390/d11030046
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.