The wonderful world of the yeast Saccharomyces cerevisiae...a glimpse!
Dr. Fahd Nasr
Professor in Functional Genomics and Translational Biology, Dean, Academic Strategist, Curricula Developer, Academic Counselor, Freelancer, Life Coach
A huge toolbox to approach basic eukaryotic molecular mechanism
It is the most intensively studied eukaryotic model organisms in molecular and cell biology, much like Escherichia coli as the model prokaryote. The yeast S. cerevisiae is a very tractable model organism to study other models systems including multicellular and complex models. Many reasons favor the use of fungi in general: relatively small and haploid genome, short life cycle, easy to manipulate, sexual phase and meiosis, etc…
A model for [molecular] genetics…
Yeast is suitable for doing genetics, easy and economical growth, hundreds of interesting mutants, simple equipment, and even a pleasant smell! Genetic analysis of the yeast is undemanding: making mutants, crossing, cloning and deleting genes, transposon mutagenesis, etc. Above all, yeast is a eukaryote, fit to study a range of genetic and physiological processes. Yeast’s entire genomic sequence is available in databases.
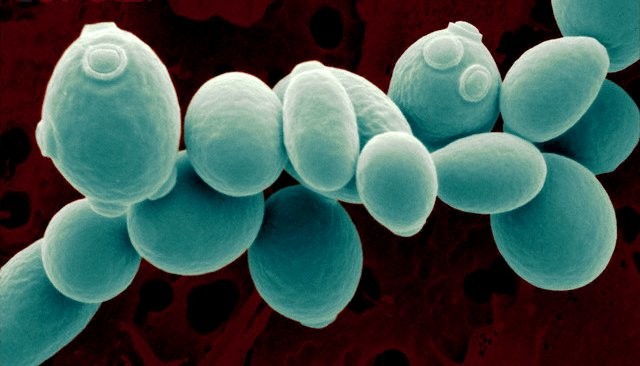
The eukaryotic cell…
A cell of the budding yeast S. cerevisiae is about 4-7micro m, bud scars are visible on cycling cells. Incomplete budding results in cells that remain attached, the chains of loosely joined cells form the pseudohyphae. However, S. cerevisiae does not produce hyphal structures as other ascomycetes. Meiosis can be controlled. Diploid cells can be induced to undergo meiosis and sporulation under nitrogen starvation or on acetate agar. Contrary to the fission yeast, budding gives two cells of unequal size: a mother, which may enter a new cell cycle, and a daughter that needs to mature first before cycling. Yeast cells age as humans… A yeast cell can’t divide for ever! Usually, cells age and mothers lose their cycling potential and die after approximately 30-40 divisions.
Called the bakers yeast…
Yeast ferments sugars and make dough rise. It is behind the most common type of fermentation. Where? It is found everywhere, on fruits, flowers and other sugar containing substrates! Yeast can tolerate a range of growth conditions: as low as freezing to about 55°C, complete drying, sugar concentrations of 3M, 20% alcohol, etc. However, yeast can proliferate from 12°C to 40°C, with low pH (2.8-8.0) and high ethanol tolerance.
It is safe…
Moreover, yeast is a safe microorganism to produce commercially important proteins. We use the term GRAS to designate the yeast S. cerevisiae. GRAS means Generally Recognized As Safe! Biotechnological production of human therapeutic agents in yeast is approved… it does not require the same extensive experimentation demanded for unapproved host cells! For a range of reasons proteins isolated from the yeast represent a better product than is obtainable using a prokaryotic expression system. Yeast can be seen as a model and a tool! A model to study the living cell, and a tool to improve existing (or to generate new) biotechnological processes.
Cytoduction…what a technique!
In S. cerevisiae, mating of haploid cells results in cell fusion and stable diploids, which are prototrophs with complementing auxotrophies. Fusion of the haploid nuclei or karyogamy is delayed in kar1 mating allowing kar-mediated plasmid and other cytoplasmic factors transfer between yeast strains. Cytoduction is the production of cells with mixed cytoplasm but with the nucleus of one or the other parent. We call such cells cytoductants! Cytoduction is regarded as a gene transfer technique and as a tool in studying the cytoplasmic heredity in yeast.
Let’s make cytoduction simple or not!
A cell that mates with a kar1-1 mutant cell forms a zygote with two unfused nuclei, one WT and one with the kar1-1 allele…then, the first bud from the zygote contains only one parental haploid nucleus, and the cytoplasmic material from both parental strains. In this kar1 mediated gene transfer heterokaryons with mixed cytoplasm are produced, and DNA molecules can be transferred. When I say DNA molecules, I refer to plasmids but also to whole chromosomes! With proper selection such transfer is made directional. Controlled cytoduction is based on kar1 mating; it enables the directional transfer of material from donor cells into recipient cells. The haploid cells resulting from cytoduction are called cytoductants, chromoductants, and plasmoductants. They differ in properties. Cytoductants refer to haploid progeny obtained from a kar mating; they have the nucleus of the recipient and mixed cytoplasm from both parents. As to chromoductants and plasmoductants, they are haploid progeny with the nucleus of the recipient and a chromosome or plasmid from donor, respectively. In the cytoduction scheme, excon-jugants bearing the nucleus of one parent and the cytoplasmic markers of the other can also be produced!
A really tractable system!
The facility of yeast manipulation is astounding! Nitrogen starvation triggers meiosis and sporulation to form an ascus with four haploid spores. It might seem a very simple microorganism, however, the yeast can exist in different cell types with different genetic programs! In the yeast S. cerevisiae one can distinguish haploids MATa vs MATalpha, haploid vs diploid, mother vs daughter, and spores! Segregation of chromosomes and markers can be followed by tetrad analysis; the four ascospores in each ascus are the products of meiosis. Such packaging of the four meiotic products makes genetic analysis particularly simple in yeast.
Yeast cells shmoo…to meet!
The yeast cells can be maintained either as haploid or diploid, a property that makes the yeast very attractive to geneticists. Mating in yeast is easy to explain: haploid cells produce mating peptide pheromones, i.e. a-factor and alpha-factor, to which the mating partner responds to prepare for mating. Two haploid cells can mate to form a zygote 2n; since yeast cannot move, cells must grow towards each other (cytoplasmic extensions are called shmoos). Pheromone response is a signal transduction pathway; it is central to sexual communication and controls the response of yeast cells to a- or alpha-factor. The pathway consists of a specific pheromone receptor that binds a- or alpha-factor; Binding of pheromone stimulates reorientation of the cell towards the source of the pheromone (the mating partners).
Enough literature…get into molecular aspects!
Pheromone binding stimulates the MAP signaling cascade that causes cell cycle arrest to prepare cells for mating (cell synchronization in G1 phase). The pheromone MAP signaling cascade is a sophisticated pathway! Well, let’s start with the receptors of the alpha and a pheromones, which are Ste2 or Ste3 GPCR, respectively. Following pheromone binding, a heterotrimeric G protein (Gpa1/Scg1 Gα , Ste4 Gß, and Ste18 Gγ) is activated. Then, Gßγ dimer binds the Ste5 scaffold protein and activates a MAPK pathway composed of three players: Ste11 MAPKKK, Ste7 MAPKK, and Fus3 MAPK. Note that in the absence of pheromone signaling, Gpa1 binds to the Ste4-Ste18 Gßγ dimer to prevent activation of the MAPK pathway. There is an alternative pathway for the activation of the MAPK pathway involving the same Ste4-Ste18 Gßγ dimer. In this alternative pathway, Ste4-Ste18 Gßγ dimer binds Ste20 MAP4K that Phosphorylates and activates the Ste11 MAPKKK!

We call it the iceberg concept…
It is clear that genes coding for the receptors are the only required to distinguish between a and alpha haploid cells! Downstream of the receptors pheromone interaction the cascade of events of the MAP pheromone response pathway is the same in both cells! It has been shown that mutations that eliminate steps downstream of the receptor/pheromone interaction have the same effects on both cell types! Only haploid cells of opposite sex can mate to form a diploid a/alpha. These cells of different sex have different genetics programs!
A startling feature…switch at will!
Haploid (n) and diploid (2n) cells do have different genetic programs. For instance, n-specific genes code for proteins involved in the response to pheromone.What determines the sex type of a haploid cell is a locus dubbed MAT on chromosome III of the yeast genome. A startling feature of yeast cells is their ability to switch their mating type, from a to alpha and vice versa; switching occurs once per cell division!!! For example, an a cell will switch to an α cell by replacing the MATa allele with the MATα allele. How does this switching happen? In fact copies of MATa and MATα alleles are present but only in a silenced state. They are located on chromosome III like the active MAT locus! We call them silent mating cassettes HML, an inactive copy of the MATα allele, and HMR, a silenced copy of the MATa allele.

Nothing magic! It is about recombination…
HML and HMR are the abbreviations of “Hidden MAT Left” and “Hidden MAT Right” loci on chromosome III, respectively. Well, only MAT locus is active. To become active the information present in the silent cassettes must be 'read into' the MAT locus by gene conversion. Gene conversion is an event in DNA genetic recombination resulting in a nonreciprocal transfer of genetic information. Yes, that is why the replacement of MAT active sequence using either HML or HMR information is a gene conversion process. This is a mechanistically very difficult issue. As to the genetic programs that control the behavior of haploid cells, several regulatory proteins are encoded by the mating type locus. The MATa locus encodes the a1 transcriptional activator and MATalpha locus encodes the alpha1 activator and the alpha2 repressor. In short, alpha1 activator stimulates alpha-specific genes and the alpha2 repressor represses a-specific genes in alphas cells! In diploid cells the a1/alpha2 heteromeric repressor prevents the expression of all haploid-specific genes including the famous RME, a repressor of meiosis.
This post is intended to simply introduce to you the fascinating world of the yeast Saccharomyces cerevisiae; thanks to more than 100 years of extensive investigation, the yeast system boasts of uniques features for all scientists regardless of their research interests.
Fine regards to all.
Prof. Fahd Nasr
#yeast #Saccharomycescerevisiae #genetics #matingtype #modelorganism #genomics #lebaneseuniversity #profnasr
The following webpage is mainly dedicated to Yeasts used as models and tools:
https://yeastwonderfulworld.wordpress.com/