
Parrotfish grazing ability: interspecific differences in relation to jaw-lever mechanics and relative weight of adductor mandibulae on an Okinawan coral reef
- Published
- Accepted
- Received
- Academic Editor
- John Ringo
- Subject Areas
- Ecology, Marine Biology, Zoology
- Keywords
- Parrotfish, Chlorurus, Scarus, Grazing ability, Interspecific difference, Excavator, Scraper, Jaw-lever mechanics
- Copyright
- © 2016 Nanami
- Licence
- This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, reproduction and adaptation in any medium and for any purpose provided that it is properly attributed. For attribution, the original author(s), title, publication source (PeerJ) and either DOI or URL of the article must be cited.
- Cite this article
- 2016. Parrotfish grazing ability: interspecific differences in relation to jaw-lever mechanics and relative weight of adductor mandibulae on an Okinawan coral reef. PeerJ 4:e2425 https://doi.org/10.7717/peerj.2425
Abstract
Parrotfishes (family Labridae: Scarini) are regarded to have important roles for maintaining the ecosystem balance in coral reefs due to their removal of organic matter and calcic substrates by grazing. The purpose of the present study was to clarify the interspecific differences in grazing ability of five parrotfish species (Chlorurus sordidus, C. bowersi, Scarus rivulatus, S. niger and S. forsteni) in relation to interspecific differences in jaw-lever mechanics and the relative weight of the adductor mandibulae (muscles operating jaw closing). The grazing ability was calculated by using stomach contents (CaCO3 weight/organic matter weight) defined as the grazing ability index (GAI). There were significant interspecific differences in GAI (C. sordidus = C. bowersi > S. rivulatus > S. niger = S. forsteni). Teeth of C. sordidus and C. bowersi were protrusive-shape whereas teeth of S. rivulatus, S. niger and S. forsteni were flat-shape. C. sordidus and C. bowersihave jaw-lever mechanics producing a greater biting force and have a larger weight of adductor mandibulae. S. rivulatus has jaw-lever mechanics producing a greater biting force but a smaller weight of adductor mandibulae that produce an intermediate biting force. In contrast, S. niger and S. forsteni have jaw-lever mechanics producing a lesser biting force and have a smaller weight of adductor mandibulae. Feeding rates and foray size of S. rivulatus, S. niger and S. forsteni were greater than C. sordidus and C. bowersi. The degree in bioerosion (GAI × feeding rate) was the largest for S. rivulatusand the smallest for S. forsteni. The degree in bioerosion for C. sordidus was larger than S. niger whereas relatively equal between C. bowersi and S. niger. These results suggest that interspecific difference in GAI was explained by interspecific differences in teeth shape, jaw-lever mechanics and relative weight of adductor mandibulae. The interspecific difference in the degree of bioerosion suggests the importance of various size of parrotfishes with diverse feeding modes to maintain healthy coral reef ecosystems.
Introduction
Coral reefs support high species diversity of marine organisms including fishes. Among the diverse species of coral reef fishes, parrotfishes (family Labridae: Scarini) are considered to be important components to maintain a healthy coral reef ecosystem in relation to their feeding mode (reviewed in Bonaldo, Hoey & Bellwood, 2014). Parrotfishes are considered to contribute to the enhancement of settlement of benthic organisms including corals by removing epilithic algae and benthic organisms on the substrates by gazing (Mumby et al., 2006; Adams et al., 2011 but also see Russ et al., 2015). Parrotfishes are also considered to contribute to the bioerosion of coral reefs which is an important process to build coral reef environments (Bellwood, 1995a; Bellwood, 1995b). A decrease in parrotfish density due to human activities would subsequently cause dramatic change in the ecosystem balance and resilience of coral reefs (Bellwood et al., 2004; Bellwood, Hoey & Hughes, 2011; Hughes et al., 2007).
Bellwood & Choat (1990) categorized parrotfishes into two functional groups based on the morphological characteristics of jaws (premaxilla, maxilla, dentary and articular). The first group is formed of excavators that have a deep shape and a thick cement covering of jaws. Feeding behavior of excavators shows a short powerful bite and the degree of grazing was greater, which was found by their bite marks termed as scars (Bellwood, 1995a; Bellwood, 1995b; Bellwood, 1996; Alwany, Thaler & Stachowitsch, 2009; Bonaldo & Bellwood, 2009). Excavators mainly consist of genus Bolbometopon, Cetoscarus, Chlorurus and some Sparisoma species (Bellwood & Choat, 1990; Bellwood, 1994). The second group is formed of scrapers that have a shallow shape and thin cement covering of jaws. Scrapers deliver weaker bites and graze less, per bite unit, than excavators (Lokrantz et al., 2008; Bonaldo, Krajewski & Bellwood, 2011). Scrapers mainly consist of the genus Scarus and several Sparisoma species (Bellwood & Choat, 1990; Bellwood, 1994).
Some previous studies have estimated the grazing ability of parrotfishes by observation of scars (Bellwood, 1995a; Bellwood, 1995b; Bellwood, 1996; Van Rooij, Videler & Bruggemann, 1998; Alwany, Thaler & Stachowitsch, 2009; Bonaldo & Bellwood, 2009; Bonaldo, Krajewski & Bellwood, 2011). Scars are found when parrotfishes graze the substrates for feeding and the characteristics of the scars were different between excavators and scrapers in relation to their jaw morphology (e.g., Alwany, Thaler & Stachowitsch, 2009; Bonaldo & Bellwood, 2008; Bonaldo & Bellwood, 2009; Lokrantz et al., 2008; Bonaldo, Krajewski & Bellwood, 2011). These studies measured the size of scars (depth × width × length) for quantitative estimations of the grazing ability and showed that the size of scars was greater for excavators than scrapers (Hoey & Bellwood, 2008; Lokrantz et al., 2008; Bonaldo & Bellwood, 2009). Several studies have also shown that the larger size of scars was found for larger size of individuals, indicating the ontogenetic difference in grazing ability of parrotfishes (Bruggemann et al., 1996; Bonaldo & Bellwood, 2008; Lokrantz et al., 2008).
Bruggemann et al. (1996) also estimated the grazing ability of two parrotfish species based on the stomach contents and showed that the grazing ability of excavator species was greater than scraper species. Ontogenetic differences in grazing ability were also found for the two species: the smaller-sized individuals (less than 14 cm in fork length) did not graze on the carbonate substrates and removed only epilithic algae, whereas larger-sized individuals (over 15 cm in fork length) ingested the calcium carbonate substrates during grazing events. Thus, Bruggemann et al. (1996) also showed that the feeding mode and body size are good indicators of the degree of grazing ability of parrotfishes.
Although parrotfishes are major fish assemblage components in Okinawan coral reefs, ecological studies on grazing and bioerosion focusing on interspecific differences in jaw-lever mechanics and relative weight of adductor mandibulae (muscle operating jaw closing) have not been clarified yet in this region (Bonaldo, Hoey & Bellwood, 2014). In addition, although jaw-lever mechanics and relative weight of adductor mandibulae are the key aspects to clarify the feeding ecology of marine fishes, quantitative comparisons of interspecific differences in jaw-lever mechanics and relative weight of adductor mandibulae have not been sufficiently examined for multiple species of parrotfishes.
The aims of the present study were to investigate the grazing ability of five species of parrotfishes in an Okinawan coral reef. Specifically, the aims were to clarify (1) interspecific differences in grazing ability, (2) ontogenetic variations in grazing ability, (3) a quantitative description of jaw-lever mechanics, (4) an interspecific comparison in relative adductor mandibulae weight to parrotfish size and (5) the feeding behavior of parrotfishes in this region.
Materials and Methods
The study was conducted mainly using field observations of free-living fishes in their natural habitat. Individuals caught for sampling were immediately killed by placing them on ice to minimize pain. The sampling procedure was approved by Okinawa prefectural government fisheries coordination regulation No. 41, which permits capture of marine fishes on Okinawan coral reefs for scientific purposes.
Study species
Five parrotfish species were selected for the present study (Chlorurus sordidus, C. bowersi, Scarus rivulatus, S. niger and S. forsteni) (Fig. 1). These five species are commonly found in Okinawa.

Figure 1: Photographs of the five parrotfish species with their upper jaw lever-system (premaxilla + maxilla) and lower jaw lever system (dentary + articular).
{kind=link}
Estimation of grazing ability using stomach contents
The grazing ability of parrotfish was estimated from stomach contents as follows. Since the substrate of coral reefs consists of limestone, the stomach contents of parrotfish can be considered to contain the calcium carbonate (CaCO3). Namely, the stomach contents can be regarded as mixtures of organic matter (e.g., epilithic algae) and CaCO3 (stomach contents = organic matter + CaCO3) (Fig. 2). In the present study, it was considered that the ratio of CaCO3 in the stomach contents would be higher if the grazing ability is greater (i.e., deep biting, Fig. 2). Therefore, the ratio (CaCO3 weight/organic matter weight) is defined as the “grazing ability index” (GAI) in the present study.
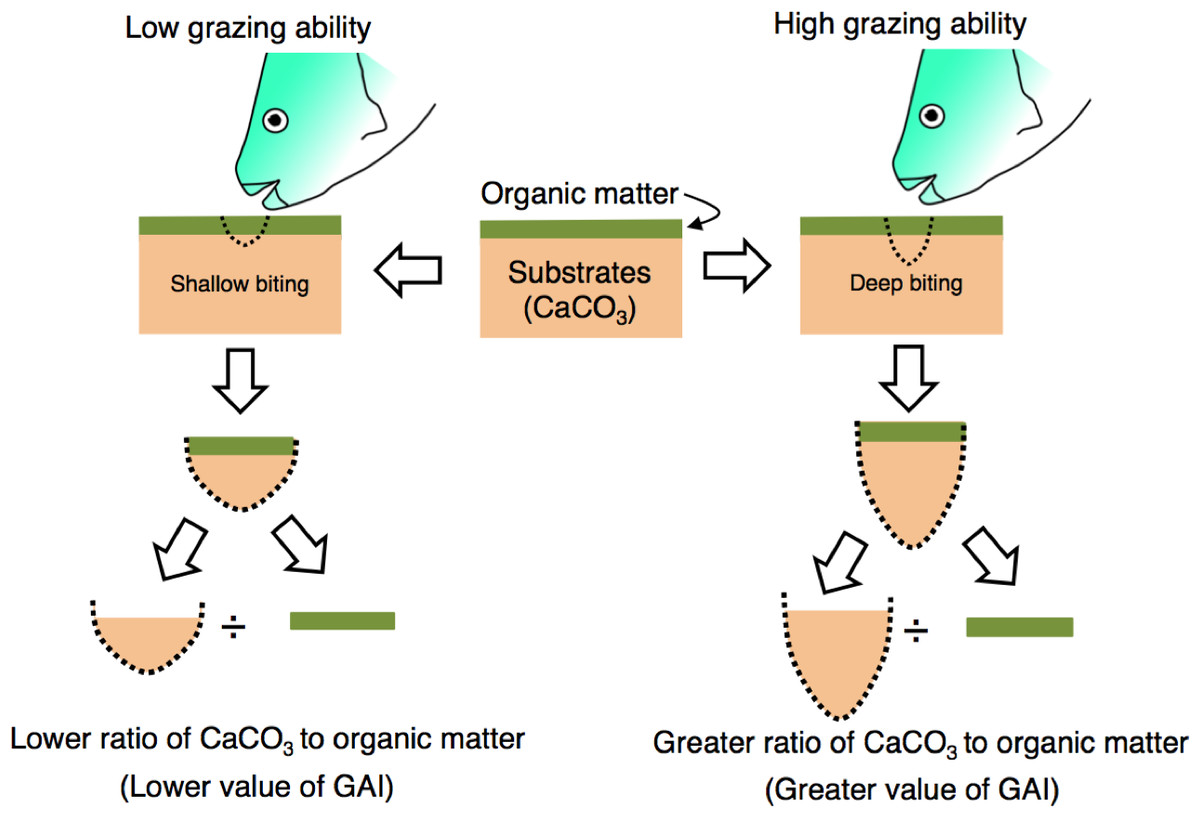
Figure 2: Schematic diagram of the definition of grazing ability index (GAI).
Substrates consist of calcium carbonate (CaCO3) and are covered by organic matter (epilithic algae, sensu Bellwood & Choat, 1990). The ratio of CaCO3 weight to organic matter weight would change with the difference in grazing ability. For measurement procedures of the CaCO3 weight and organic matter weight, see ‘Materials and Methods’.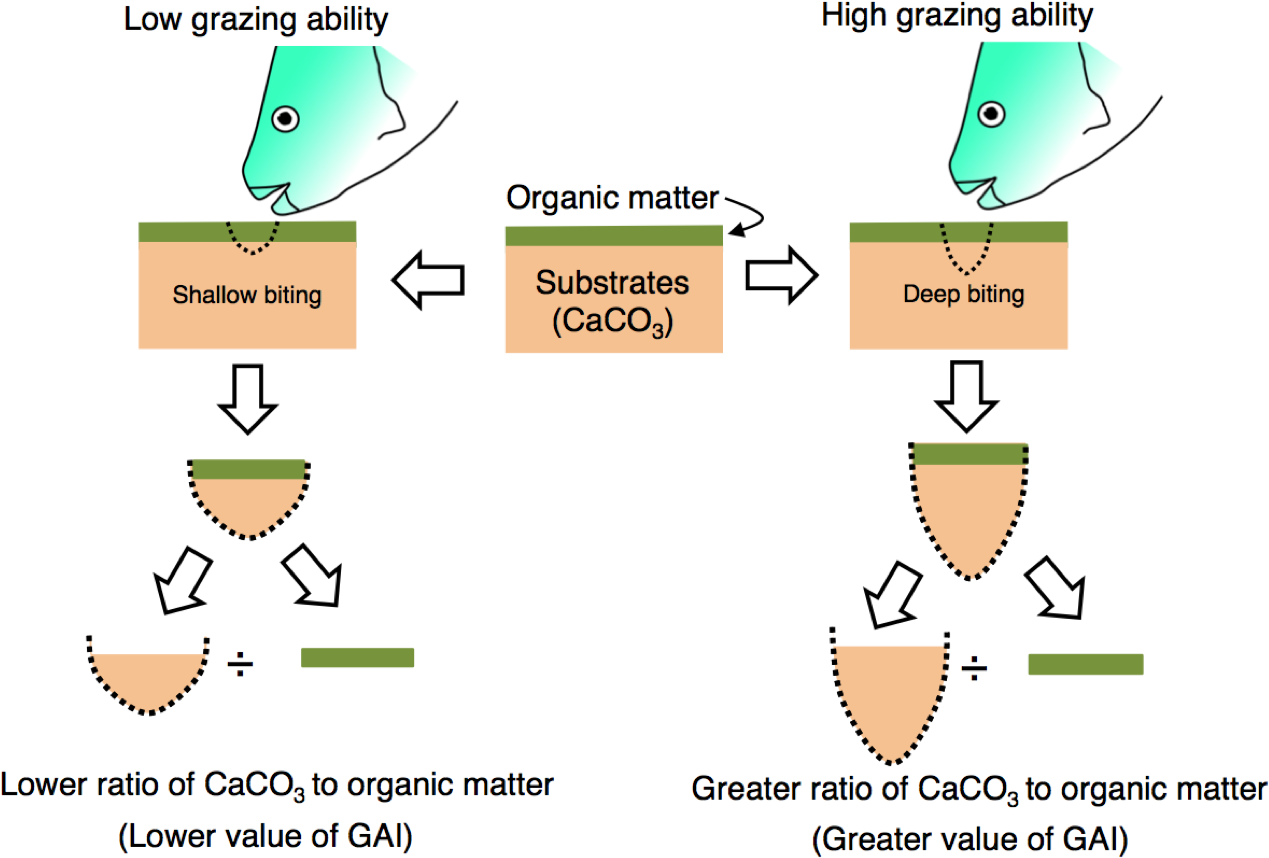{kind=link}
The CaCO3 weight was calculated as: CaCO3 can be removed from the stomach contents by adding hydrochloric acid (HCl) aqueous solution as the following reaction formula: where CaCl2 is calcium chloride, H2O is water and CO2 is carbon dioxide. Since CaCl2 is water-soluble and CO2 isvolatile, the CaCO3 as well as CaCl2 and CO2 were completely removed from the stomach contents by water rinsing. Then, the ratio of CaCO3 to organic matter weight (i.e., GAI) was calculated as: Thus, the GAI represents the CaCO3 weight that was simultaneously grazed per unit of weight of organic matter on substrate.
Sampling and experimental procedure
For the GAI estimation, five parrotfish species were sampled by spear gun between July 2013 and February 2016 from the reef slope at Ishigaki Island, Okinawa, Japan. The number of individuals and the range of fork length (FL) were as follows: C. sordidus (n = 33: 149.5–236.0 mm), C. bowersi (n = 23: 179.5–265.5 mm), S. rivulatus (n = 26: 154.0 mm–319.5 mm), S. niger (n = 20: 166.0–288.0 mm) and S. forsteni (n = 22: 145.0–303.5 mm). All fishes were collected by spearing the head and immediately killed in an icebox to minimize any loss of stomach contents.
Dry weight in the stomach contents and organic matter were determined as follows: (1) a subsample of stomach contents (c.a. 3 g in wet weight) was put into a 1.5 ml microtube and dried at 58 °C; (2) the weight of the dried stomach contents was measured to the nearest 0.0001 g; (3) 1N-HCl aqueous solution was poured onto the stomach contents; (4) the microtube was centrifuged and supernatant HCl aqueous solution was removed using micro-pipet with care not to remove any stomach contents; (5) the third and fourth procedures were repeated until no CO2 bubbles were found during the third procedure; (6) the remaining stomach contents were rinsed using distilled water, centrifuged and the supernatant water was removed. Finally, the organic matter (stomach contents with no CaCO3) was dried at 58 °C and the weight was measured to the nearest 0.0001 g.
Interspecific comparison in GAI
The relationships between FL and GAI were plotted as a logarithmic function as: where a and b are coefficients. One-way ANOVA was conducted for interspecific comparisons of GAI for four FL ranges with 40-mm intervals: (1) 140.0 mm–180.0 mm, (2) 180.5 mm–220.0 mm, (3) 220.5 mm–260.0 mm and (4) over 260.5 mm. If a significant difference was found, a post-hoc Games-Howell test was applied for multiple comparisons among the five species.
Teeth characteristics
Number of teeth on dental plates for premaxilla and dentary was counted. A total of 10 samples were used for each species. One-way ANOVA and post-hoc Games-Howell test was used for significant differences in the ratio among the five species.
Jaw-lever mechanics for closing
Jaw-lever mechanics for closing was compared by measuring the morphological characteristics of the upper jaw (premaxilla + maxilla) and lower jaw (dentary + articular) in accordance with Wainwright & Bellwood (2002), Westneat (2004) and Westneat et al. (2005). Jaw-lever mechanics are related to the force and velocity in the feeding of fishes (see Westneat, 2004; Westneat et al., 2005). In accordance with Bellwood & Choat (1990), the jaw-lever mechanics for closing was investigated (Fig. S1). The length of in-lever-closing (Lin-lever-closing) and out-lever (Lout-lever) were measured by using a digital caliper (Fig. 3). Then, the ratio of in-lever-closing to out-lever (Lin-lever-closing/Lout-lever) were calculated and compared among the five species. The greater values in the ratio means greater biting force but lower velocity in biting and vice versa (Westneat, 2004). A total of 10 samples were used for each species. One-way ANOVA and post-hoc Games-Howell test was used to test for significant differences in the ratio among the five species.
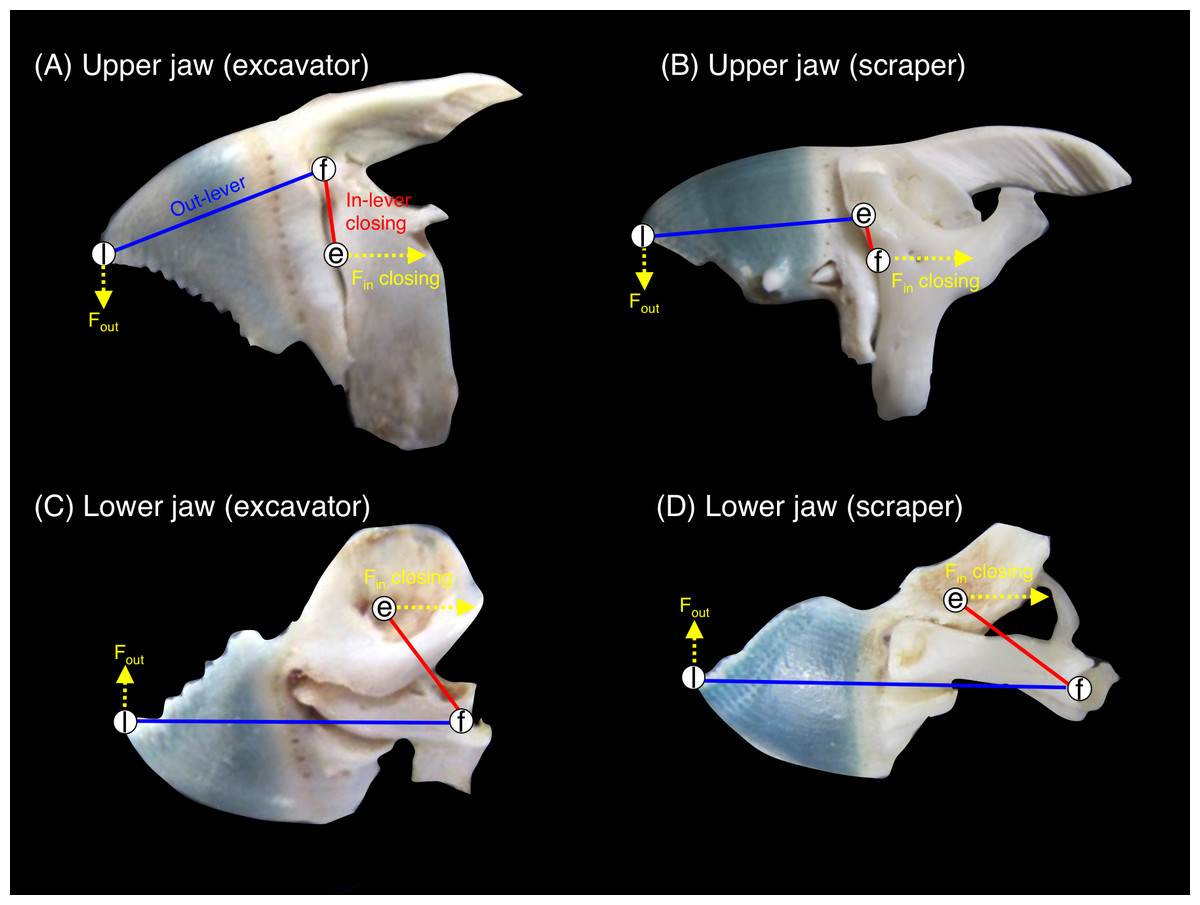
Figure 3: Pictorial explanation of the jaw-lever mechanics for closing (e, effort; f, fulcrum; l, load. See also Fig. S1).
(A) upper jaw of excavator (Chlorurus sordidus); (B) upper jaw of scraper (Scarus forsteni); (C) lower jaw of excavator (Chlorurus sordidus); (D) lower jaw of scraper (Scarus forsteni). The relative difference in biting force and velocity can be estimated by measuring the length of the in-lever-closing (red lines: Lin-lever-closing) and out-lever (blue lines: Lout-lever). The ratio of in-lever-closing to out-lever (Lin-lever-closing/ Lout-lever) can be calculated in accordance with Wainwright & Bellwood (2002), Westneat (2004) and Westneat et al. (2005). The greater values in the ratio means greater biting force but lower velocity in biting and vice versa. Fin-closing: force for jaw closing at tendon (see also Fig. S1). Fout: produced force for jaw rotation.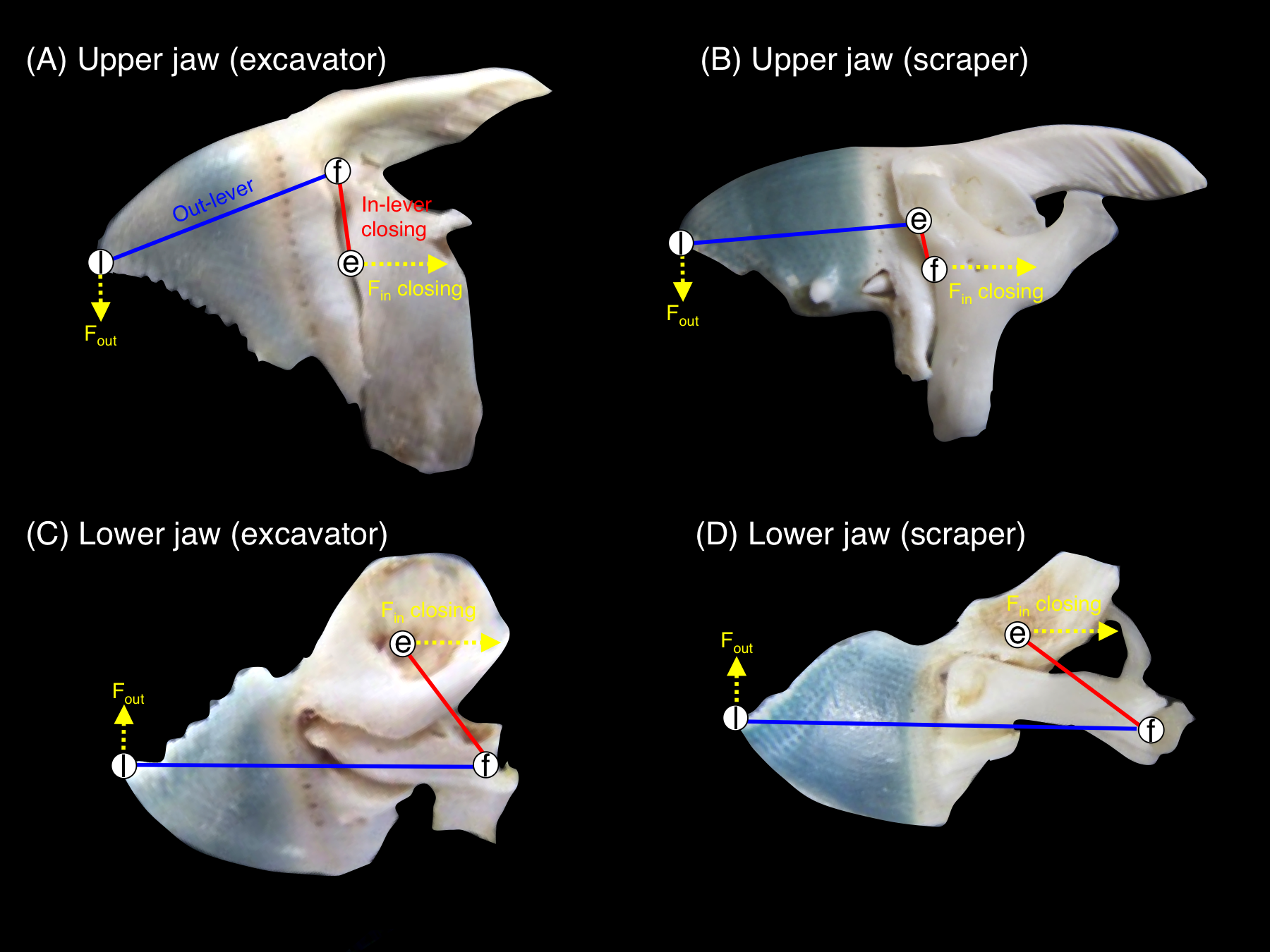{kind=link}
Muscles operating the jaw
Weight of muscles operating jaw (adductor mandibulae) was measured nearest 0.01 g by dissecting. The relationship between FL and adductor mandibulae weight was plotted as a power function as: where a and b are coefficients. Analysis of covariance (ANCOVA) was performed to compare the inter-specific difference in the FL- adductor mandibulae weight relationship. Prior to the analysis, the data was log-transformed and the relationship was converted as a simple linear regression. If a significant difference was found, a post-hoc Bonferroni test was applied for multiple comparisons among the five species.
Underwater observations of feeding behavior
Feeding behavior was observed at the fringing reef around Ishigaki Island between October and November 2011 and between October 2012 and February 2013. Underwater observations were conducted at the reef slope (fore reef) using SCUBA equipment or snorkeling. Depth range was 3 m–5 m and water temperature was 22 °C–28 °C. A researcher (A Nanami) searched for individuals of the focal species and followed a focal individual paying particular attention not to disturb the behavior of the individual. To avoid double-counts for the focal individual, species, size and/or sex were changed for the subsequent observations. The estimated FL (cm), number of bites per 5 min (feeding rate) and foray size (mean number of bites: sensu Bellwood & Choat, 1990) were counted. The FL range was between 25 cm and 35 cm and time zone was between 1200 h–1600 h. Since previous studies have demonstrated a time-zone difference in feeding rates (e.g., 0800 h–1000 h, 1001 h–1200 h, 1201 h–1400 h and 1401 h–1600 h; Bellwood, 1995a; Bonaldo & Bellwood, 2008), a one-way ANOVA was performed to clarify if any time-zone differences occurred in the feeding rates for the two time zones (1200 h–1400 h and 1401 h–1600 h). As a result, no significant time-zone differences were found for all five species. Thus, the feeding rates obtained in the two time zones were pooled and used for further analyses. The numbers of individuals were 10, 11, 11, 7 and 8 for Chlorurus sordidus, C. bowersi, Scarus rivulatus, S. niger and S. forsteni, respectively. One-way ANOVA and a post-hoc Games-Howell test were applied for multiple comparison for feeding rates and foray size among the five species.
During the observations of feeding behavior, the numbers of bites per substrate were also recorded. Previous studies have categorized the feeding substrates as epilithic algae on hard substrata, macroalgae, live coral, crustose coralline algae and seagrass (Rotjan & Lewis, 2005; Rotjan & Lewis, 2009; Hoey & Bellwood, 2008; Alwany, Thaler & Stachowitsch, 2009; Nash et al., 2012; Bonaldo, Hoey & Bellwood, 2014). The data for the respecting feeding substrates were represented as the proportion of the number of bites (%) for each substrate against the total number of bites. The proportions of bites for each individual were averaged for each species.
Interspecific comparison in degree of bioerosion
From the data of GAI and feeding rates, the degree of bioerosion was calculated as follows:
This was because the degree of bioerosion should be considered by both GAI and feeding rates. For calculation of the degree of bioerosion, calculated GAI data at 30 cm FL were obtained using the above-mentioned FL-GAI relationship. The rate of focal species to another species was calculated for all pairs of the five species for interspecific comparison of the degree of bioerosion.
Results
Interspecific difference in GAI
There were significant positive relationships between FL and GAI for four species (Chlorurus sordidus, C. bowersi, Scarus rivulatus and S. niger) (Fig. 4). Although a slight positive relationship between FL and GAI was found for S. forsteni, it was not significant (Fig. 4). Overall, the GAIs were greater for C. sordidus and C. bowersi, intermediate for S. rivulatus and lesser for S. niger and S. forsteni (Fig. 5). For the 141 mm–180 mm FL range, the GAI of C. sordidus was significantly greater than S. niger and S. forsteni (Games-Howell test, p < 0.05) (Fig. 5A). For the 181–220 mm FL range, GAIs of C. sordidus and C. bowersi were significantly greater than the other three species (Fig. 5B). The GAI of S. rivulatus was also significantly greater than S. niger and S. forsteni (Fig. 5B). For the 221 mm–260 mm FL range, the GAIs for C. sordidus, C. bowersi and S. rivulatus were significantly greater than S. niger and S. forsteni (Fig. 5C). For the over 260 mm FL range, significant differences in GAI were found among S. rivulatus, S. niger and S. forsteni (Fig. 5D).
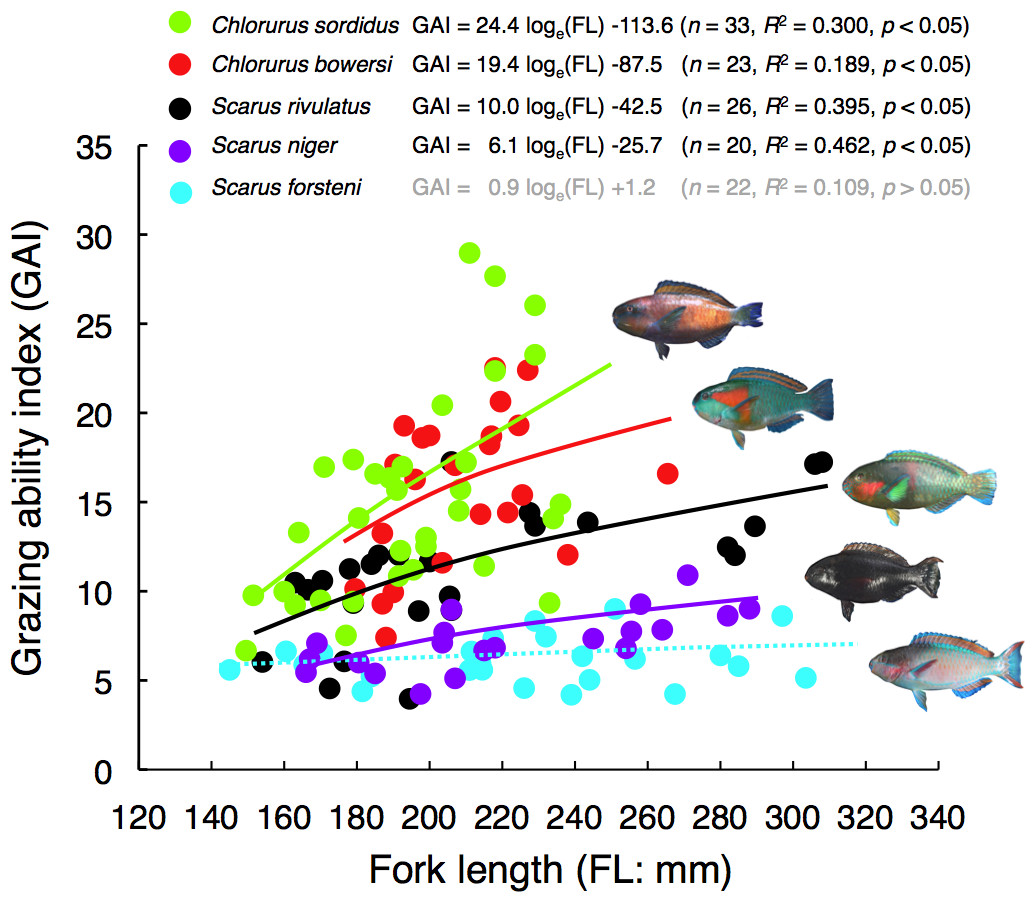
Figure 4: Relationships between fork length (FL) and grazing ability index (GAI) for the five parrotfish species.
The relationships are plotted by logarithmic function.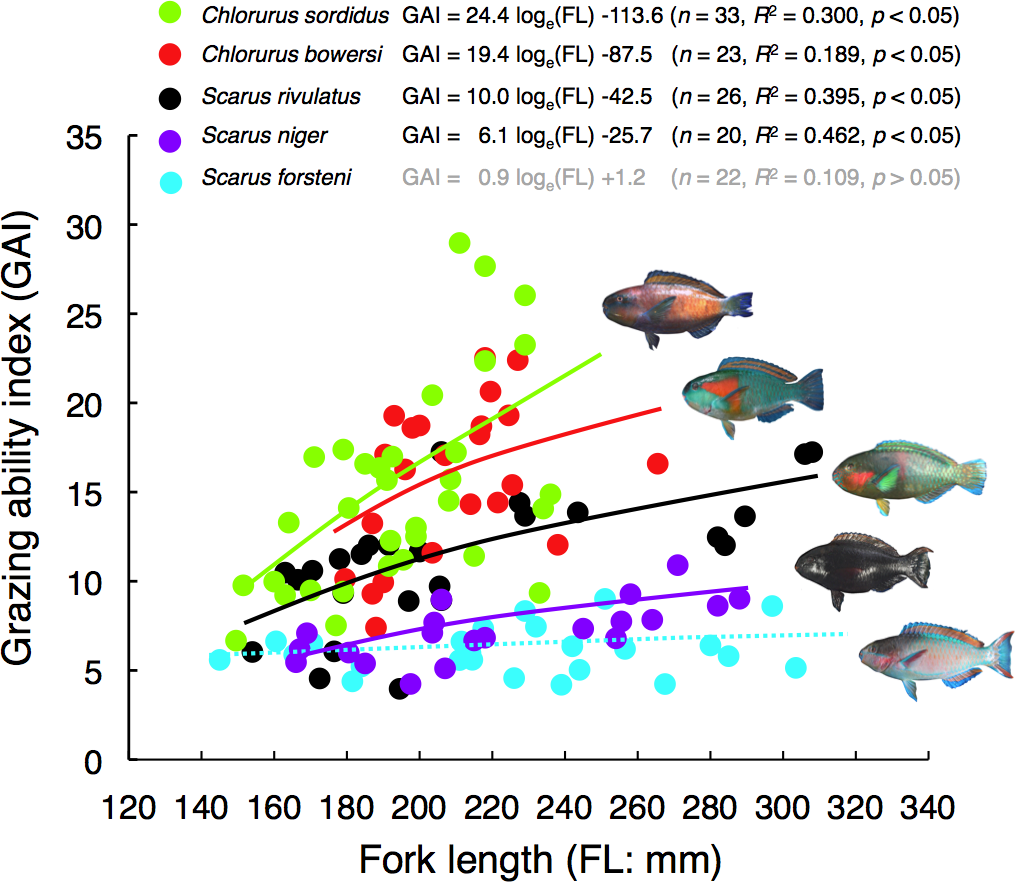{kind=link}
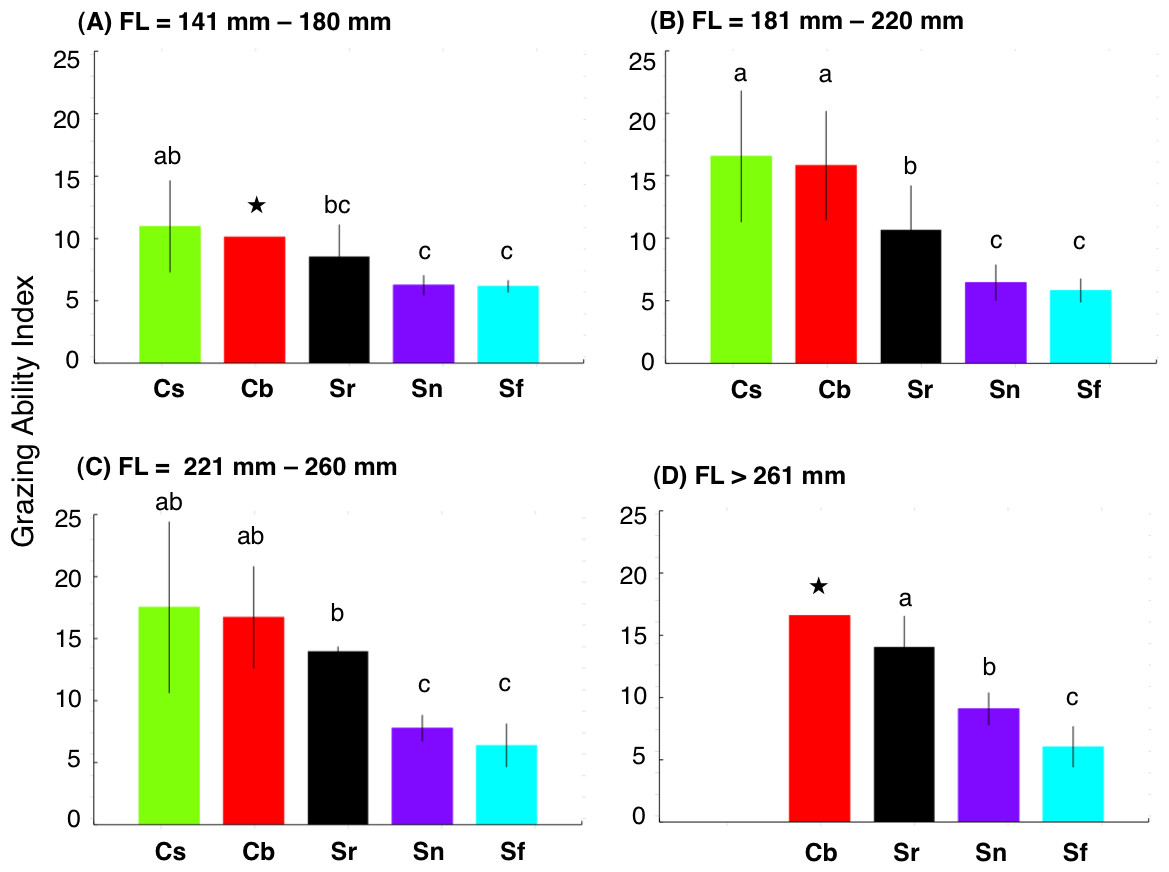
Figure 5: Interspecific difference in grazing ability index (GAI) for four fork length (FL) ranges.
The different alphabet characters on the bars indicate the significant difference in GAI (Games-Howell test, p < 0.05). Vertical lines represent standard deviation. Star symbols in (A) and (D): only 1 individual for Chlorurus bowersi and excluded from Games-Howell test due to no standard deviations. No individuals for C. sordidus in (d). Species names are abbreviated as Cs, Chlorurus sordidus; Cb, C. bowersi; Sr, Scarus rivulatus; Sn, S. niger; Sf, S. forsteni.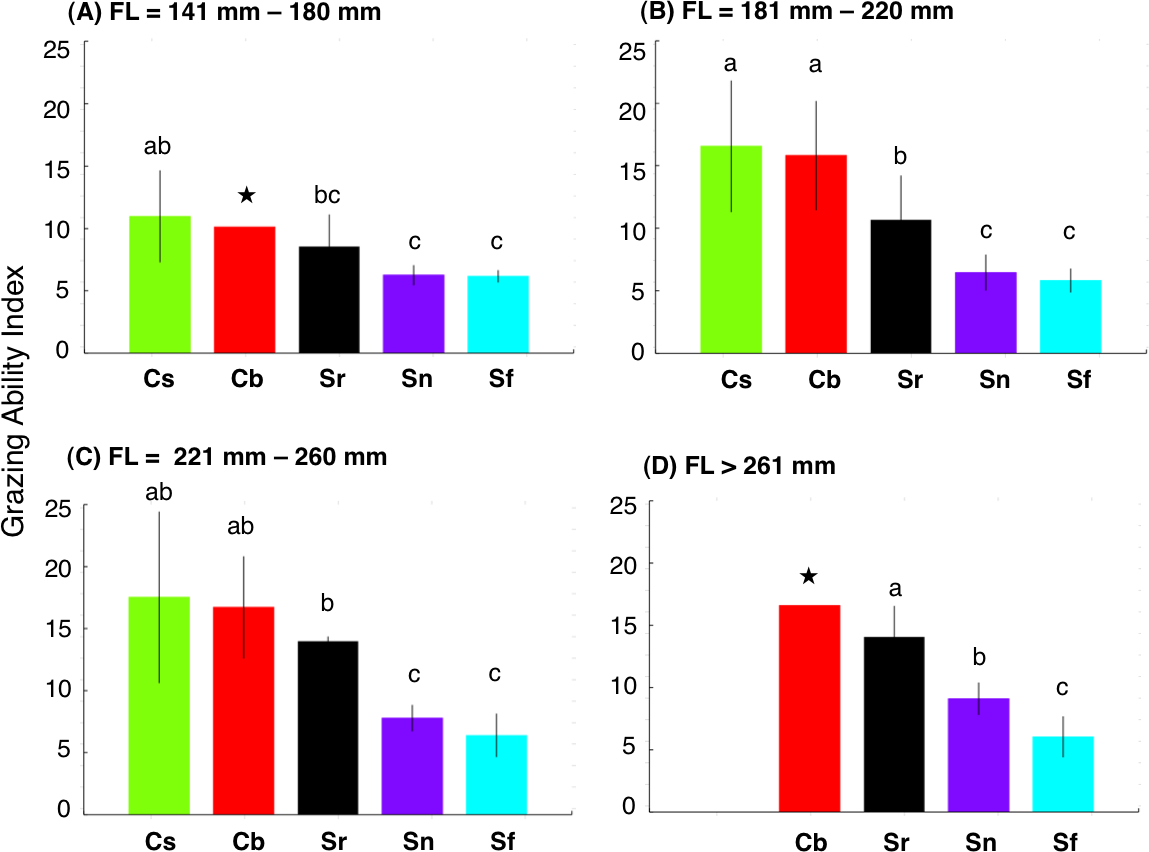{kind=link}
Interspecific differences in teeth characteristics
Each tooth was protrusive-shape for C. sordidus and C. bowersi whereas each tooth was relatively flat-shape for S. rivulatus, S. niger and S. forsteni for premaxilla (Fig. 6A) and dentary (Fig. 6B). The number of teeth of premaxilla for C. sordidus, S. rivulatus and S. forsteni was significantly greater than C. bowersi and S. niger (Fig. 6C). The number of teeth of dentary for S. rivulatus and S. forsteni was significantly greater than C. sordidus, C. bowersi and S. niger (Fig. 6D).
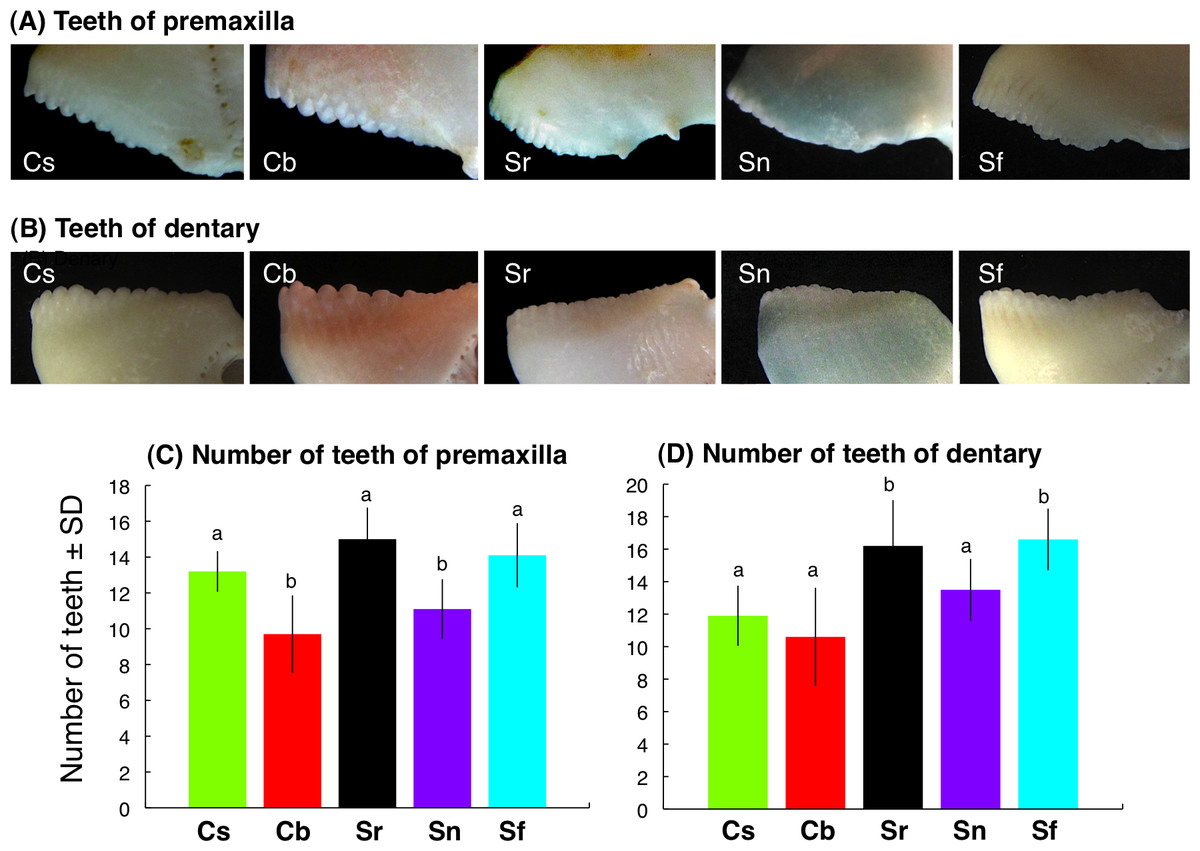
{kind=link}
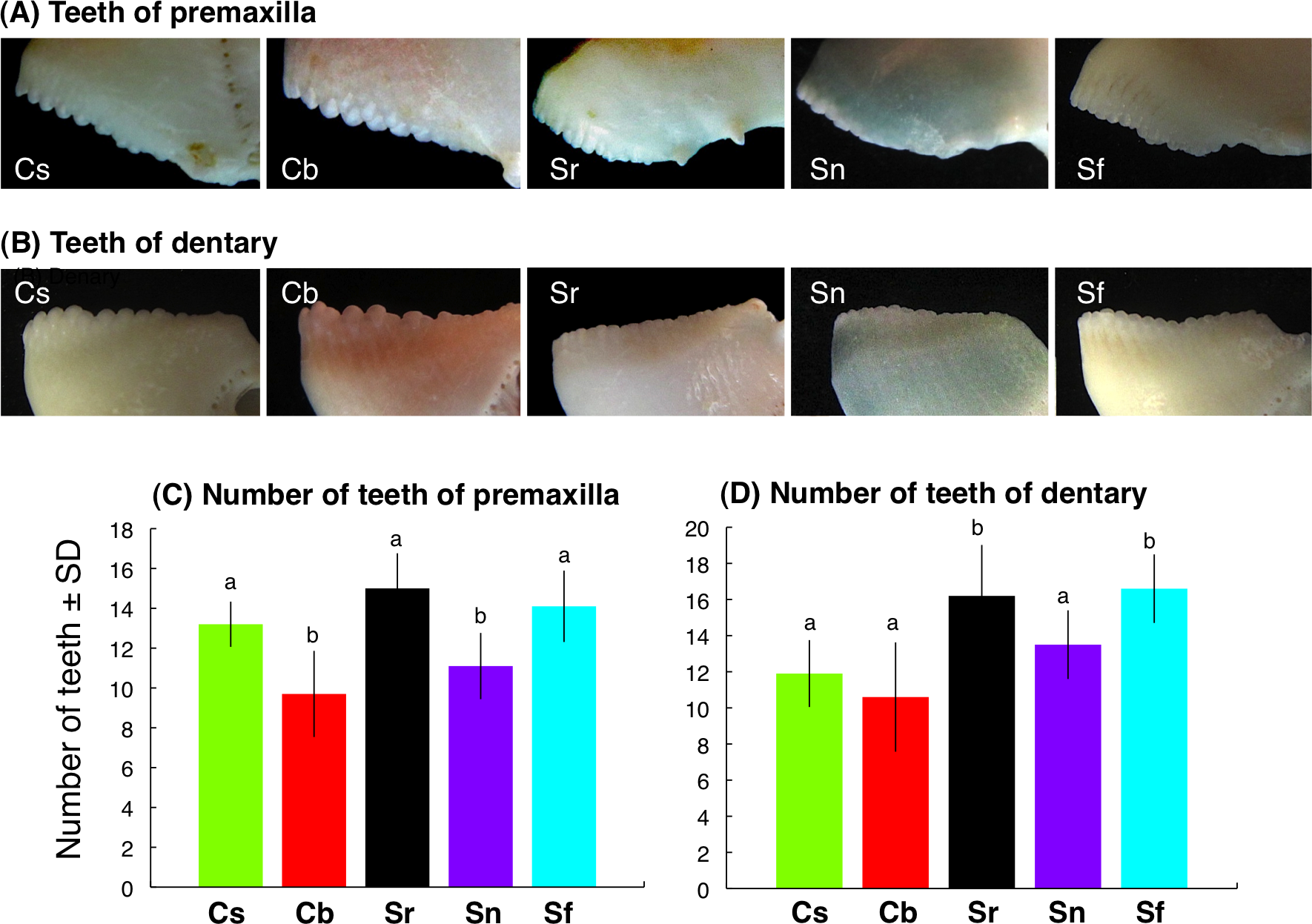
Interspecific differences in jaw-lever mechanics for closing
For the upper jaw, the ratio of in-lever-closing to out-lever (Lin-lever-closing/Lout) was significantly different among the five species (C. sordidus > C. bowersi > S. rivulatus > S. niger > S. forsteni) (Fig. 7A) (one-way ANOVA and post-hoc Games-Howell test, p < 0.05). For the lower jaw, the ratio of in-lever-closing to out-lever (Lin-lever-closing/Lout) for C. sordidus and C. bowersi was significantly greater than S. niger and S. forsteni (Fig. 7B). No significant difference in the ratio was found among C. sordidus, C. bowersi and S. rivulatus and between S. rivulatus and S. niger (Fig. 7B).
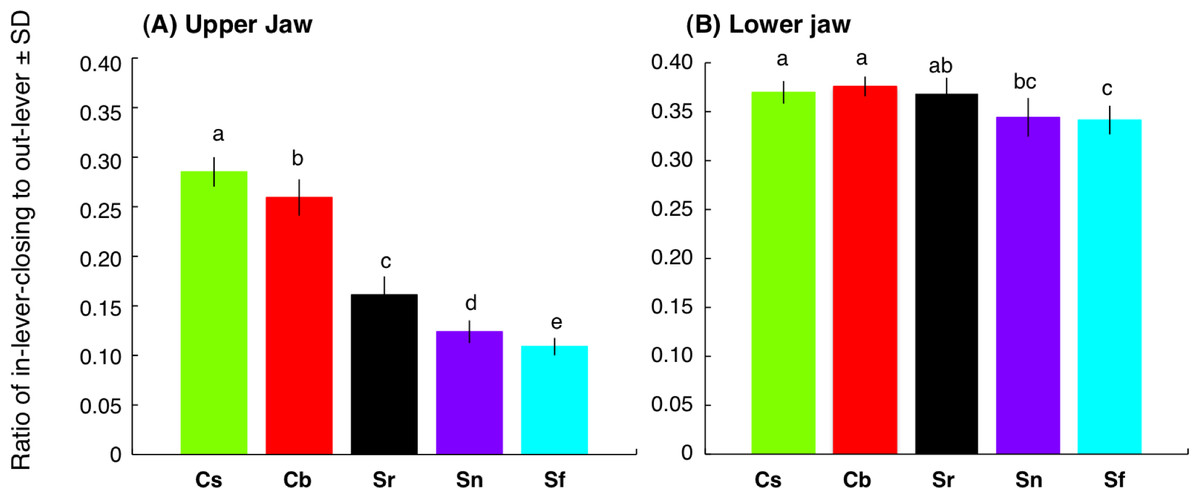
Figure 7: Interspecific difference in the ratio of in-lever-closing to out-lever (Lin-lever-closing/ Lout-lever) (see also Fig. 3) for upper jaw (A) and lower jaw (B).
The greater values in the ratio means higher force but lower velocity, whereas lower values in the ratio means higher velocity but lower force (Westneat, 2004). Different alphabet characters on the bars indicate significant difference (one-way ANOVA and post-hoc Games- Howell test, p < 0.05). Species names are abbreviated as Cs, Chlorurus sordidus; Cb, C. bowersi; Sr, Scarus rivulatus; Sn, S. niger; Sf, S. forsteni.{kind=link}
Interspecific difference in relationship between FL and adductor mandibulae weight
Relative weight of adductor mandibulae against FL for C. sordidus and C. bowersi was significantly greater than the other three species (ANCOVA and post-hoc Bonferroni test, p < 0.05) (Fig. 8). The relative weight for C. sordidus was significantly greater than C. bowersi. In contrast, no significant difference was found among S. rivulatus, S. niger and S. forsteni (ANCOVA and post-hoc Bonferroni test, p > 0.05) (Fig. 8).
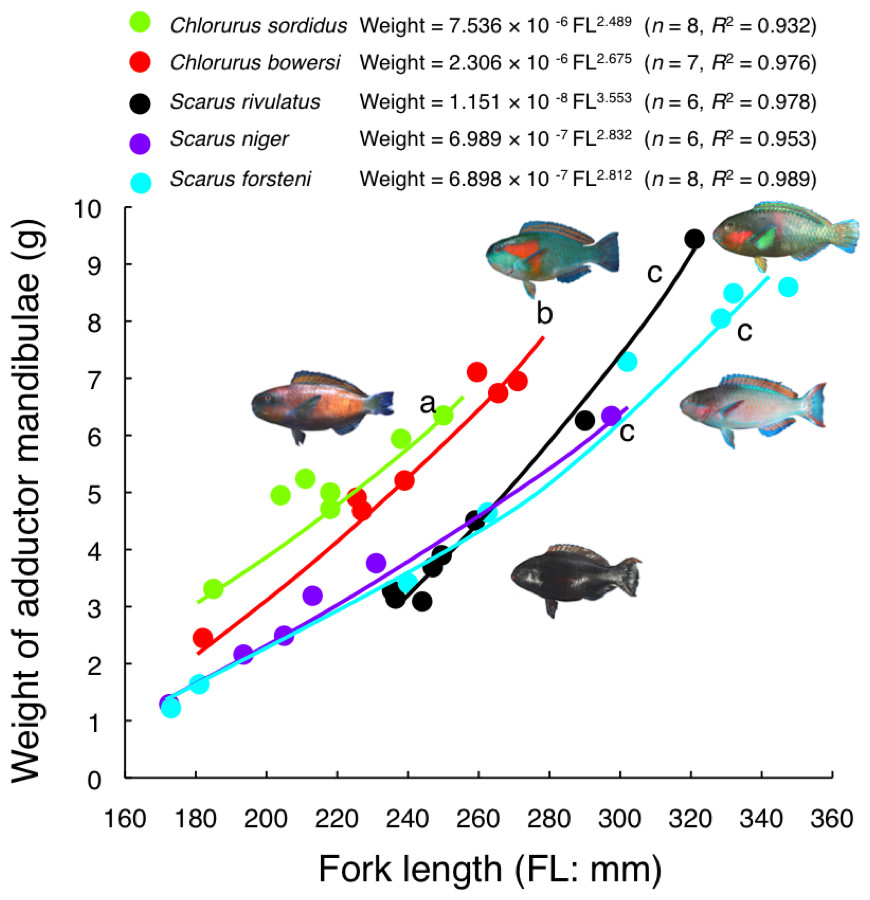
Figure 8: Relationship between fork length and weight of adductor mandibulae for the five parrotfish species.
Different alphabet characters near the regression lines indicate significant difference (ANCOVA and post-hoc Bonferroni test, p < 0.05).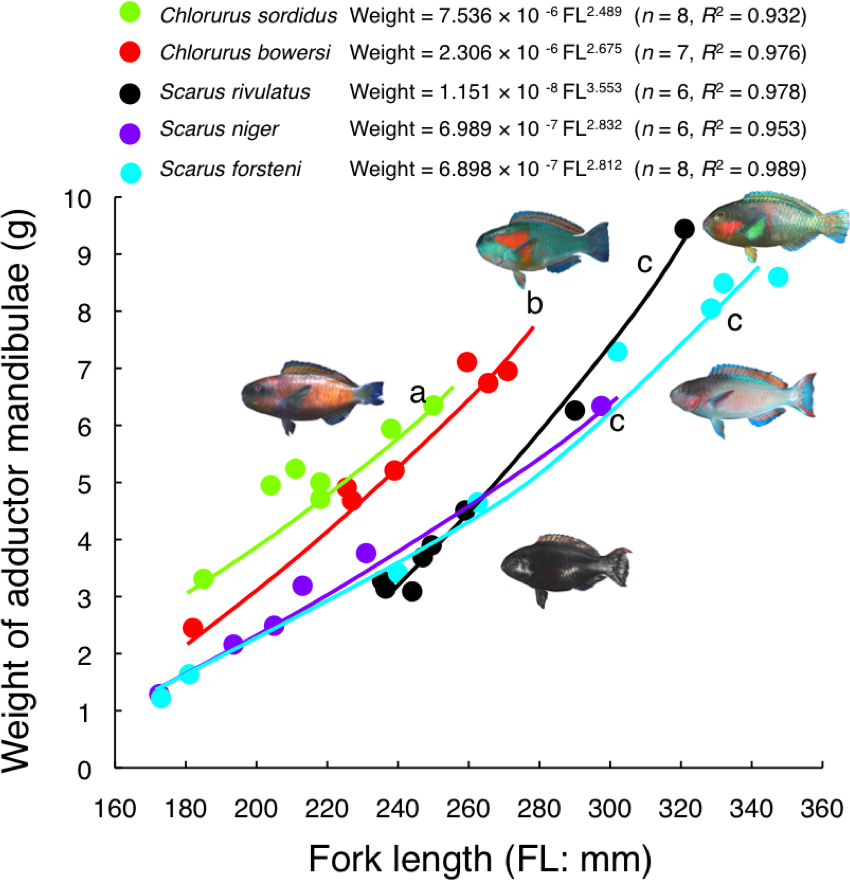{kind=link}
Feeding behavior
Feeding rates for Scarus rivulatus, S. niger and S. forsteni tended to be higher than C. sordidus and C. bowersi (Table 1). The feeding rates of S. rivulatus and S. niger were significantly higher than C. sordidus and C. bowersi (one-way ANOVA and post-hoc Games-Howell test, p < 0.05). A similar tendency was found for foray size (Table 1). The foray size of S. rivulatus and S. niger were significantly larger than C. sordidus and C. bowersi (one-way ANOVA and post-hoc Games-Howell test, p < 0.05). The main feeding substrate was epilithic algae on hard substrates such as rocks and dead coral skeletons. No individuals were observed feeding on macroalgae, crustose coralline algae or seagrass in the present study. Live coral were rarely fed on by C. bowersi and S. niger (Table 1). C. sordidus, S. rivulatus and S. forsteni did not feed on live coral.
| Species | n | Feeding rates (5 min−1) | Foray size | Proportion of bites (%) | |
|---|---|---|---|---|---|
| ±SD | ±SD | Epilithic algae | Live coral | ||
| Chlorurus sordidus | 10 | 75.0 ± 31.7ab | 6.3 ± 2.1ac | 100 | 0 |
| C. bowersi | 11 | 66.3 ± 29.0ab | 6.1 ± 3.5ac | 99.8 | 0.2 |
| Scarus rivulatus | 11 | 186.0 ± 50.7c | 15.8 ± 7.8b | 100 | 0 |
| S. niger | 7 | 158.0 ± 53.3cd | 15.9 ± 4.6b | 98.3 | 1.7 |
| S. forsteni | 8 | 111.8 ± 37.9bd | 9.2 ± 4.7bc | 100 | 0 |
Notes:
- SD
-
Standard deviation
Interspecific comparison in degree of bioerosion
The degree of bioerosion for S. rivulatus was the greatest (1.41–2.66 times greater than the other four species) and the smallest for S. forsteni (0.38–0.71 times greater than the other four species) (Table 2). For the other three species (C. sordidus, C. bowersi and S. niger), degree of bioerosion for C. sordidus was greater or for S. niger (1.25 times greater) whereas the degree of bioerosion was almost equal between C. bowersi and S. niger. The differences in degree of bioerosion were relatively similar among C. sordidus, C. bowersi and S. niger (0.75 times–1.19 times greater for each species comparison).
| Focal species | Compared species | GAI (times) | Feeding rate (times) | Degree in bioerosion (GAI × Feeding rate) (times) |
|---|---|---|---|---|
| Chlorurus sordidus | C. bowersi | 1.10 | 1.13 | 1.25 |
| S. rivulatus | 1.76 | 0.40 | 0.71 | |
| S. niger | 2.81 | 0.47 | 1.33 | |
| S. forsteni | 2.81 | 0.67 | 1.89 | |
| C. bowersi | C. sordidus | 0.91 | 0.88 | 0.80 |
| S. rivulatus | 1.59 | 0.36 | 0.57 | |
| S. niger | 2.55 | 0.42 | 1.07 | |
| S. forsteni | 2.55 | 0.59 | 1.51 | |
| Scarus rivulatus | C. sordidus | 0.57 | 2.48 | 1.41 |
| C. bowersi | 0.63 | 2.81 | 1.76 | |
| S. niger | 1.60 | 1.18 | 1.88 | |
| S. forsteni | 1.60 | 1.66 | 2.66 | |
| S. niger | C. sordidus | 0.36 | 2.11 | 0.75 |
| C. bowersi | 0.39 | 2.38 | 0.94 | |
| S. rivulatus | 0.63 | 0.85 | 0.53 | |
| S. forsteni | 1.00 | 1.41 | 1.41 | |
| S. forsteni | C. sordidus | 0.36 | 1.49 | 0.53 |
| C. bowersi | 0.39 | 1.69 | 0.66 | |
| S. rivulatus | 0.63 | 0.60 | 0.38 | |
| S. niger | 1.00 | 0.71 | 0.71 |
Notes:
- GAI
-
Grazing ability index
Discussion
Grazing ability index (GAI) to estimate interspecific difference in grazing ability
The present study is the first trial to design a new index of parrotfish grazing ability using stomach contents and to estimate interspecific differences in grazing ability among multiple parrotfish species using the new index. The GAI can be considered as an indicator to enable comparison of the interspecific difference in grazing ability among multiple parrotfish species.
Interspecific difference in GAI in relation to jaw-lever mechanics
This study is the first study to estimate parrotfish grazing ability in an Okinawan coral reef. Furthermore, although the grazing abilities of Chlorurus sordidus, Scarus rivulatus and S. niger were estimated by previous studies in the Great Barrier Reef and the Red Sea (e.g., Lokrantz et al., 2008; Bonaldo & Bellwood, 2008; Alwany, Thaler & Stachowitsch, 2009), the grazing abilities for C. bowersi and S. forsteni are first estimated in the present study. The present study also showed the degree of increase in GAI with FL was different among the species. It is suggested that the interspecific differences in GAI among the five species was explained by the interspecific differences in jaw-lever mechanics and relative weight of the adductor mandibulae. Two excavator species (C. sordidus and C. bowersi) have protrusive-shape teeth and jaw-lever mechanics that produce a greater biting force for both upper and lower jaws than the other three species (S. rivulatus, S. niger and S. forsteni). These two species also showed greater weight of adductor mandibulae, contributing to produce a greater biting force. As a result, the GAI would increase with the increase in FL. In contrast, two scraper species (S. niger and S. forsteni) have flat-shape teeth and jaw-lever mechanics producing a lower biting force for both upper and lower jaws as well as smaller weight of adductor mandibulae. Consequently, GAI of the two species would be smaller than the other three species (C. sordidus, C. bowersi and S. rivulatus) and GAI would not dramatically increase despite the increase in FL. The present study firstly demonstrated that GAI of S. rivulatus was greater than the other two scraper species (S. niger and S. forsteni), although S. rivulatus is categorized into scrapers (Bellwood & Choat, 1990). S. rivulatus has jaw-lever mechanics for upper jaws that produces the larger biting force among the three scraper species. Although jaw-lever mechanics for the lower jaw was not significantly different among C. sordidus, C. bowersi and S. rivulatus, the relative weight of adductor mandibulae was significantly less among these three species. Thus, it is suggested that S. rivulatus produce an intermediate biting force among the five species. Since the genus Scarus consists of 18 species in Okinawa and have various morphological characteristics (Nakabo, 2000), interspecific differences in GAI in further multiple species should be conducted in the future study.
Interspecific comparison in feeding behavior and degree of bioerosion
Feeding rate and foray size for S. rivulatus and S. niger was significantly greater than C. sordidus and C. bowersi. This tendency was consistent with other previous studies indicating that the feeding rates and foray size for scrapers was greater than excavators (reviewed in Bonaldo, Hoey & Bellwood, 2014). Among the scrapers, the feeding rates and foray size for S. forsteni was smaller than the other two species. The present study estimated the interspecific difference in degree of bioerosion using GAI and feeding rate. Overall, the degree of bioerosion for S. rivulatus was the greatest and the smallest for S. forsteni. The degree of bioerosion of C. sordidus was greater than for S. niger whereas almost the same between C. bowersi and S. niger. These results suggest that although the GAI was lower for scrapers, scrapers showed greater feeding rates and as a consequence, degree of bioerosion would be greater than excavators in some cases (e.g., S. rivulatus in the present study).
The results of the present study suggest the importance of various sizes of parrotfishes with diverse feeding modes. Smaller-sized individuals would contribute to the removal of organic matter (e.g., epilithic algae) on the substrates in a less destructive manner. In contrast, larger-sized individuals, especially for excavators, would contribute to the removal of both organic matter and calcic substrates in a more destructive manner. In some cases such as S. rivulatus, which has an intermediate GAI and higher feeding rate, would more effective contribution to bioerosion. Since it has been suggested that increased fishing pressure leads to a decrease in the mean size of parrotfishes (Vallès & Oxenford, 2014; Vallès, Gill & Oxenford, 2015), effective protection of various sizes as well as the various species of parrotfish on reefs should be conducted to maintain a healthy ecosystem balance and resilience in coral reefs.
Supplemental Information
Jaw-lever mechanics of parrotfishes
Observation was based on Bellwood & Choat (1990). An example for Chlorurus sordidus is shown. In (A), yellow arrows show the pulling direction of tendons for jaw closing. In (B), blue and red letters (fulcrum, effort and load) represent the three kinetic points for upper jaw and lower jaw, respectively.
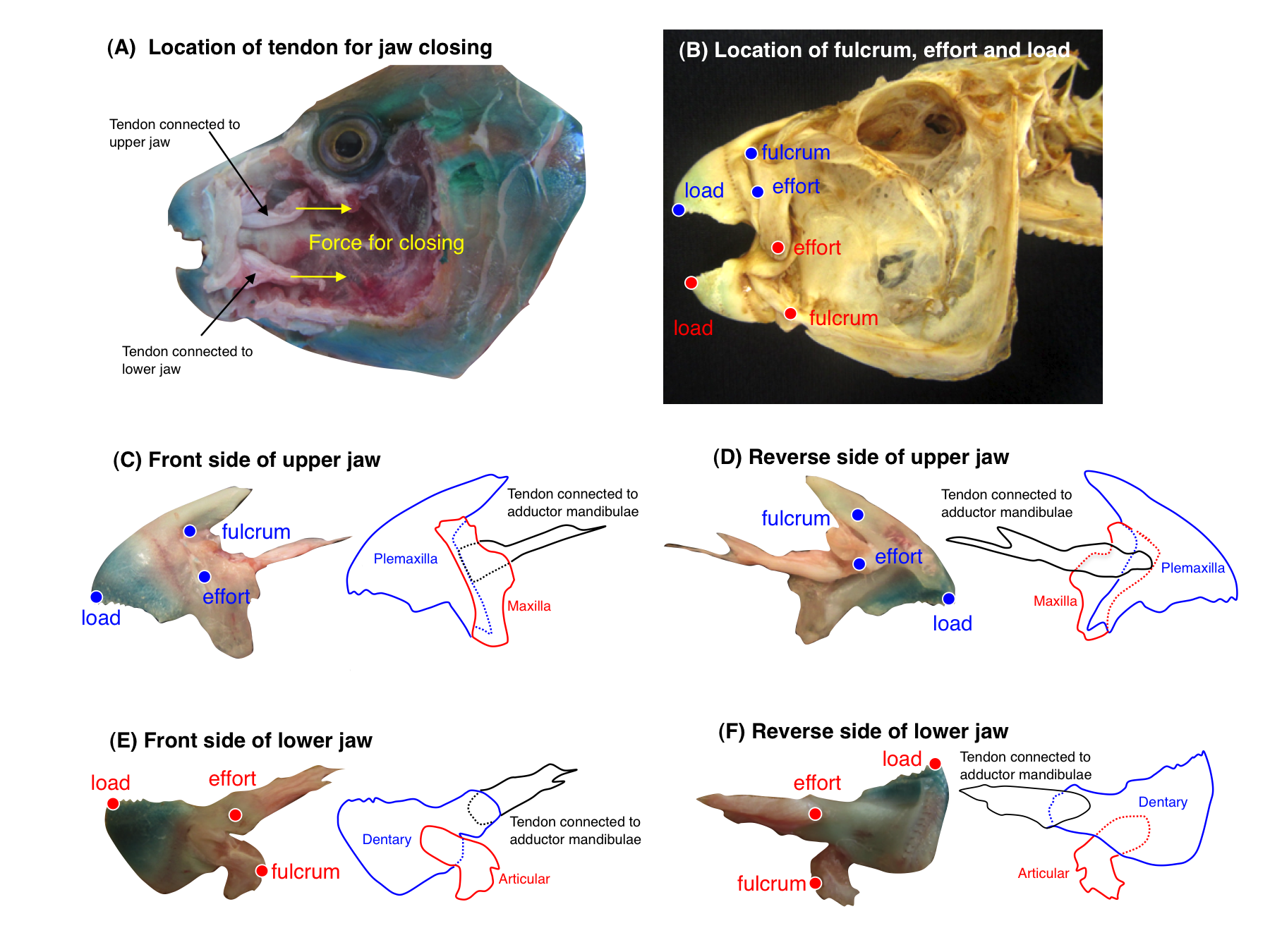{kind=link}